


ABSTRACT
Ticks and tick-borne diseases (TBD) continue to pose a significant threat to livestock production and rural livelihoods in sub-Saharan Africa (SSA). The control of ticks has relied predominantly on the use of chemical acaricides; however, the widespread and often indiscriminate application of these compounds has led to the rapid emergence and spread of acaricide resistance. This review provides a comprehensive synthesis of current knowledge on acaricide resistance in both single-host ticks, such as
Keywords: acaricide resistance, biological control, climate change, integrated tick management, multi-host ticks, single-host ticks, sub-Saharan Africa, tick-borne diseases.
INTRODUCTION
Ticks and tick-borne diseases (TBD) remain a major constraint to livestock production in sub-Saharan Africa (SSA), threatening both animal health and rural livelihoods. As obligate hematophagous ectoparasites, ticks transmit a range of protozoal, bacterial, and viral pathogens responsible for economically devastating diseases, including East Coast fever (
The epidemiology of TBD in SSA is strongly shaped by ecological and climatic factors. Warm, humid environments such as the Great Rift Valley and coastal zones support year-round tick activity, whereas climate change is expanding tick habitats into previously marginal areas and higher altitudes, intensifying disease risk [5]. Tick population dynamics are further influenced by livestock density, vegetation type, and seasonal rainfall, which together determine the frequency of acaricide use and the likelihood of acaricide resistance development [6]. Over the past decades, chemical acaricides, including synthetic pyrethroids, organophosphates, and amidines, have remained the cornerstone of tick control. However, the heavy and often indiscriminate application of these compounds has driven widespread resistance in key vectors such as
Resistance evolution is particularly pronounced in single-host ticks, whose life cycles on a single animal expose them continuously to chemical treatments, thereby accelerating selection pressure. In contrast, multi-host ticks such as
The accelerating spread of acaricide resistance threatens not only livestock productivity but also the efficacy of current tick-borne disease control programs. The growing evidence of climate-driven tick range expansion, coupled with intensifying selection pressure from chemical control, underscores the urgent need for sustainable, region-specific management strategies. Integrated tick management (ITM), combining acaricide rotation, biological control, resistant livestock breeds, and environmental interventions, offers a promising alternative but remains underutilized across SSA [4, 11].
This review provides an updated synthesis of acaricide resistance in both single- and multi-host ticks in tropical and subtropical regions of SSA. We explore the mechanisms of resistance, regional patterns, and environmental drivers, identify critical knowledge gaps, and highlight opportunities for implementing ITM strategies to safeguard livestock production and rural livelihoods in a changing climate.
REVIEW METHODOLOGY
A comprehensive literature search was conducted using electronic databases including PubMed, Scopus, Web of Science, Google Scholar, and African Journals Online. The search covered peer-reviewed articles, reports, and dissertations published between 2010 and 2025 using combinations of the following keywords: “acaricide resistance”, “Rhipicephalus microplus”, “Rhipicephalus decoloratus”, “Amblyomma variegatum”, “Rhipicephalus appendiculatus”, “single-host ticks”, “multi-host ticks”, “sub-Saharan Africa”, “tick-borne diseases”, and “integrated tick management”. Additional relevant publications were identified through manual screening of reference lists from retrieved articles. Only studies reporting acaricide resistance data in cattle ticks from tropical and subtropical regions of SSA were included. Non-English language articles, conference abstracts without full text, and studies focused exclusively on non-cattle hosts were excluded. Data extraction focused on resistance status, mechanisms, geographic distribution, environmental drivers, and control strategies. A total of 45 relevant sources were synthesized narratively to provide an updated overview of the current status, drivers, and sustainable control options.
GEOGRAPHICAL PATTERNS OF ACARICIDE RESISTANCE IN SINGLE-HOST AND MULTI-HOST TICKS
In SSA, the geographical distribution of acaricide resistance among single-host ticks (
Multi-host ticks show somewhat different patterns: in East and West Africa,
DISTRIBUTION AND PATTERNS OF ACARICIDE RESISTANCE
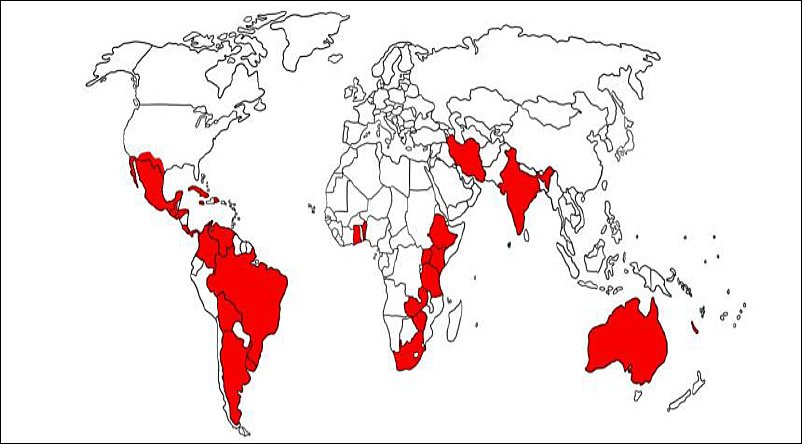
Figure 1. Global geographic distribution of acaricide resistance of single-host ticks [6].
Multiple-host ticks, including
Regional variability in resistance among multi-host ticks appears related to climatic and ecological conditions. For instance,
INFLUENCE OF ENVIRONMENTAL CONDITIONS ON ACARICIDE RESISTANCE
Environmental conditions, including climate, vegetation, and altitude, play a crucial role in the geographical distribution of acaricide resistance among tick populations. Temperature, humidity, vegetation, and altitude collectively shape tick survival, acaricide use frequency, and resistance evolution, with climate change expanding habitats into higher altitudes. Temperature and humidity have a direct influence on tick survival, reproduction, and activity, which, in turn, impact the frequency of acaricide application. For instance, high temperatures, moisture, and seasonal rainfall in East Africa and parts of Southern Africa promote tick activity and necessitate frequent acaricide use, contributing to resistance buildup in both single- and multi-host tick species [15].
Vegetation types also affect tick habitat suitability, influencing species distribution and resistance patterns. Adult
Altitude also influences tick species distribution and acaricide resistance. Higher-altitude regions, like those found in parts of Kenya and Ethiopia, are generally less favorable for ticks due to lower temperatures, lower humidity, host availability and vegetation types; however, climate change has been linked to an altitudinal range shift for ticks, increasing the likelihood of resistance spreading into previously unaffected areas [10]. These shifting geographical boundaries indicate that environmental conditions and changing climate are dynamic drivers of acaricide resistance.
IMPLICATIONS FOR REGION-SPECIFIC TICK MANAGEMENT
The regional variability in acaricide resistance necessitates tailored tick control approaches to account for local environmental conditions and tick species compositions. Climate factors, seasonality and land use are important factors for region-specific management as they impact patterns of tick distribution and resistance. In high-density cattle regions of East and Southern Africa, rotating acaricide classes and incorporating non-chemical control methods can help mitigate resistance buildup for single-host ticks [16].
Pasture management and biological control measures can contribute to reducing tick populations across both single-host and multi-host tick species by disrupting off-host survival stages and lowering overall infestation pressure. Practices such as pasture rotation and strategic resting reduce host availability for questing ticks and can limit tick survival irrespective of host-use strategy, thereby decreasing reliance on chemical control [14]. Rajakaruna and Eremeeva [17] indicated that roughly 85% livestock farmers do not practice acaricide rotation which worsens challenge of tick resistance. In areas where multi-host ticks are prevalent, integrating pasture rotation and biological control measures, such as natural predators, can reduce tick survival across diverse hosts and limit the need for chemical interventions [4]. Study conducted by Singh
To effectively address acaricide resistance, surveillance and monitoring efforts are essential to identify resistance hotspots and tailor interventions. Mapping resistance patterns at regional level provides critical information to guide acaricide use and manage resistance. Incorporating climate modeling can further aid in predicting shifts in tick distribution, allowing for proactive management as climate shifts alter tick habitat suitability [12]. Involvement of agricultural institutions, veterinary pharmaceutical companies, non-governmental organizations and research institutions to provide technical support, awareness, training, infrastructure and implementation of ITM approaches for specific region is essential for sustainable tick control.
CHALLENGES OF ACARICIDE RESISTANCE FOR SINGLE-HOST AND MULTI-HOST TICKS
Acaricide resistance is a critical challenge in the control of tick populations, particularly in SSA, where single- and multi-host ticks affect both livestock and human health. The misuse and over-reliance on acaricides, combined with limited alternative control options, have led to widespread resistance, undermining efforts to manage TBD effectively [2, 12]. This resistance complicates management strategies for both single-host ticks, such as R. microplus, and multi-host ticks, such as
MECHANISMS AND DRIVERS OF ACARICIDE RESISTANCE
Acaricide resistance in ticks is driven by various mechanisms, including target-site mutations, metabolic detoxification, and behavioral adaptations. Single-host ticks, such as
In contrast, multi-host ticks such as
ECONOMIC AND HEALTH IMPLICATIONS OF ACARICIDE RESISTANCE
The economic impact of acaricide resistance is profound, especially for smallholder farmers in SSA, who rely heavily on livestock for income and subsistence. Acaricide resistance in single-host ticks such as
Ticks and diseases they transmit pose a major challenge to animal and human health. Crimean-Congo haemorrhagic fever is a zoonotic disease transmitted by Hyalomma spp. Resistance development remains a critical challenge in acaricide use on both single-host and multi-host ticks. Mtshali
LIMITATIONS OF CURRENT ACARICIDE-BASED CONTROL STRATEGIES
Despite their widespread use, acaricides remain increasingly constrained by issues of accessibility, affordability, and declining effectiveness, particularly among smallholder farming systems in SSA. Many small-scale farmers have limited access to a broad spectrum of acaricide classes and therefore depend heavily on a small number of readily available products. Repeated and often unregulated use of the same acaricides exerts strong selection pressure on tick populations, accelerating the development and spread of resistance. This challenge is further compounded by limited farmer awareness of appropriate dosing, application intervals, and acaricide rotation principles, which undermines treatment efficacy and promotes resistance development [22]. In the case of single-host ticks, such as
Resistance dynamics in multi-host tick species are influenced by their ability to utilize multiple hosts and diverse environments, which can reduce uniform acaricide exposure and allow resistant individuals to survive and reproduce [22]. These ticks can persist in wildlife reservoirs or off-host habitats where acaricide exposure is minimal, complicating control efforts and facilitating the spread of resistant populations across landscapes. Environmental conditions, particularly high temperatures and humidity that support year-round tick activity, further intensify the need for frequent acaricide application, increasing selective pressure and accelerating resistance development [24]. Collectively, these limitations highlight the unsustainability of reliance on chemical control alone and underscore the urgent need for ITM approaches. Such strategies should combine judicious acaricide use with farmer education, resistance monitoring, biological control options, and improved husbandry practices to ensure long term effectiveness and sustainability of tick control programs [23].
THE NEED FOR ITM APPROACHES
The rise of acaricide resistance in tick populations has led to the development of ITM strategies. These strategies combine chemical, biological, resistant cattle breeds such as Nguni, and environmental control methods to effectively manage tick populations while minimizing resistance. In SSA, where TBD significantly affect livestock health and productivity, effective implementation of ITM strategies is essential for sustainable tick management [16, 17].
ACARICIDE ROTATION AND COMBINATION THERAPY
Acaricide rotation and combination therapy are two commonly employed strategies in ITM that aim to reduce resistance by minimizing selective pressure on tick populations. Acaricide rotation involves periodically changing the class of acaricide used to prevent ticks from developing resistance to any one specific chemical. This method is particularly effective for single-host ticks, such as
Combination therapy involves the simultaneous application of two or more acaricides with distinct modes of action to target different physiological pathways in ticks [25]. By exerting multiple selective pressures at the same time, this approach reduces the probability that individual ticks will survive treatment through single resistance mechanisms, thereby slowing the development and spread of acaricide resistance [22, 26]. Evidence indicates that combination therapy can be effective against both single-host and multi-host tick species when applied judiciously. For instance, the combined use of organophosphates and synthetic pyrethroids has demonstrated improved efficacy against
BIOLOGICAL CONTROL ALTERNATIVES
Biological control methods offer a promising non-chemical approach to tick management and are increasingly relevant in ITM strategies aimed at reducing acaricide reliance. Biological control agents, including entomopathogenic fungi, parasitoids, and predators, can naturally suppress tick populations without contributing to resistance. For instance,
In East and Southern Africa, research on entomopathogenic fungi has shown that they can reduce tick populations in pastures where livestock graze. These fungi can be applied to areas frequented by ticks, such as cattle rest sites, where they persist in the soil and provide long term tick control. Unlike acaricides, fungi-based control is less likely to induce resistance because the fungi kill ticks through physical invasion rather than chemical pathways [24]. However, their effectiveness can vary based on environmental conditions, such as temperature and humidity, which affect fungal spore viability. Regions with high temperatures and low humidity may face challenges in maintaining fungal efficacy [13].
Biological control also includes introducing natural tick predators, such as chickens and certain bird species, which consume ticks from livestock pastures. On the other hand, these birds could further feed on cattle blood and wounds. While not a standalone solution, predators can significantly reduce tick populations and work synergistically with other ITM components. Moreover, some African pastoralists are reintroducing traditional practices, such as burning pastures to control tick populations, though this method has mixed ecological impacts and is not always suitable for modern grazing systems [15].
INTEGRATED PASTURE AND HABITAT MANAGEMENT IN SEDENTARY AND PASTORAL SYSTEMS
Environmental control strategies, including pasture management and habitat modification, constitute an important pillar of ITM by reducing suitable tick habitats and interrupting key stages of the tick life cycle. Practices such as rotational grazing, pasture resting, and controlled stocking densities reduce host availability for ticks and can significantly lower infestation pressure. These approaches are particularly effective against multi-host tick species such as
In sedentary and semi-intensive livestock systems, pasture-based interventions can be systematically planned and implemented. Vegetation management around grazing areas, kraals, and watering points reduces humidity and shelter for off-host tick stages, thereby limiting tick survival. In some settings, carefully managed strategic pasture burning has been shown to reduce tick populations; however, this approach requires thorough ecological assessment to avoid soil degradation, biodiversity loss, and unintended disruption of ecosystem services [1].
Despite their demonstrated effectiveness, the applicability of pasture and habitat management strategies across much of SSA is limited by the predominance of extensive pastoral and transhumant production systems [27]. Communal land tenure arrangements, seasonal livestock mobility in response to rainfall and forage availability, and limited control over grazing intensity often make pasture burning, rotational grazing, and pasture resting impractical [11, 28]. In such systems, the inability to exclude livestock from specific grazing areas for extended periods reduces the effectiveness of habitat-based tick control measures [29].
Nevertheless, adapted, context-specific interventions remain feasible in pastoral settings. Localized habitat management around high-risk areas, such as communal kraals, watering points, and livestock routes, can reduce tick densities where animal aggregation is greatest [21]. Community-based coordination among pastoralists is particularly important, as isolated efforts by individual farmers are unlikely to yield sustained reductions in tick populations [20, 28]. When combined with strategic acaricide application, improved animal husbandry, and farmer education, these targeted pasture interventions can contribute meaningfully to overall ITM outcomes [17].
Evidence from humid, tick-prone regions of East Africa, including parts of Kenya and Uganda, indicates that integrating pasture management with judicious acaricide use can reduce tick burdens and slow the development of acaricide resistance [16]. However, successful implementation depends on tailoring interventions to local ecological conditions, production systems, and socio-economic realities. Consequently, pasture and habitat management should be viewed as complementary components of ITM rather than standalone solutions, particularly in pastoral landscapes where mobility and communal resource use constrain their full application [30, 31].
EDUCATION AND FARMER PARTICIPATION
Education and farmer participation have a substantial effect on the adoption and implementation of new farming techniques. Oli
KNOWLEDGE GAPS AND FUTURE RESEARCH DIRECTIONS
The Rapid Tick Exposure Test (RaTexT®) is a new method for detecting acaricide resistance in cattle ticks. This test provides rapid results within 24 hours on the exposure of partially engorged adult ticks to a specially designed acaricide-impregnated matrix [8]. Testing and monitoring of various tick species across SSA is essential for sustainable farming, as this test has been conducted in three countries in Africa. Current knowledge gaps include insufficient studies on multi-host tick resistance, limited information on resistance among wildlife tick populations, and a need for regional monitoring programs that can track and address resistance as it emerges [12, 32].
Future research should focus on identifying alternative control strategies that can complement or replace chemical acaricides. These may include biological controls, such as entomopathogenic fungi, and acaricide-free integrated management approaches, such as habitat management and tick vaccines, which could offer more sustainable long term solutions [11]. Furthermore, monitoring and collaboration between researchers, stakeholders, industry, veterinarians, and local farmers will also be critical in advancing ITM and reducing acaricide resistance over the long term [13, 28].
CONCLUSION
Acaricide resistance has emerged as a widespread and escalating challenge across SSA, affecting both single-host and multi-host tick species and undermining the effectiveness of conventional tick control strategies. The evidence synthesized in this review demonstrates that single-host ticks, particularly
From a practical perspective, these findings highlight the urgent need to transition from sole reliance on chemical control toward ITM approaches. Strategies such as acaricide rotation, combination therapy, biological control using entomopathogenic fungi, pasture and habitat management, and farmer education can collectively reduce selection pressure and improve long term control outcomes. Strengthening farmer awareness and promoting correct application practices are especially critical in smallholder systems, where misuse of acaricides remains a key driver of resistance.
A major strength of this review lies in its integrated analysis of biological, environmental, and socio-management factors influencing acaricide resistance, providing a comprehensive understanding of resistance dynamics across diverse ecological settings. However, several limitations persist, including limited data on resistance patterns in multi-host ticks, insufficient information on the role of wildlife reservoirs, and a lack of standardized, region-wide surveillance systems.
Future research should prioritize the development of sustainable alternatives to chemical control, including vaccines, advanced biological agents, and precision-based management strategies. There is also a need for coordinated regional monitoring programs, improved diagnostic tools such as rapid resistance detection assays, and stronger collaboration among researchers, policymakers, and farming communities.
In conclusion, combating acaricide resistance in SSA requires a multidisciplinary and region-specific approach that integrates scientific innovation with practical farm-level interventions. The adoption of ITM, supported by continuous surveillance, education, and policy-driven support, is essential to ensure sustainable tick control, reduce the burden of TBDs, and safeguard livestock productivity in the face of evolving environmental and resistance challenges.
AUTHORS’ CONTRIBUTIONS
MY, NN and LQ: Identified the research topic, developed methodology, and drafted the manuscript. ZM and ZTHM: Reviewed and edited the manuscript. All authors read and approved the final manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors are thankful to Döhne Agricultural Development Institute for providing necessary facilities for review.
REFERENCES
- Kamani J, Chung PJ, Lee CC, Chung YT. In search of the vector(s) of
Babesia rossi in Nigeria: Molecular detection ofBabesia rossi DNA inRhipicephalus sanguineus sensu lato ticks collected from dogs. Exp Appl Acarol 2018;76:243-248. [Google Scholar] | [Crossref] - Mekonnen S, Bryson NR, Fourie LJ, Peter RJ, Spickett AM, Taylor RJ. Comparison of three tests to detect acaricide resistance in
Boophilus decoloratus on dairy farms in the Eastern Cape Province, South Africa. J S Afr Vet Assoc 2003;74(2):41-44. [Google Scholar] | [Crossref] - Mtshali PS, Mtshali MS. In silico and phylogenetic analyses of partial
Babesia bovis gene sequences from cattle in South Africa. BMC Vet Res 2017;13:1-10. [Google Scholar] | [Crossref] - Heylen DJA, Kumsa B, Kimbita E, Frank MN, Muhanguzi D, Jongejan F. Tick communities of cattle in smallholder rural livestock production systems in sub-Saharan Africa. Parasit Vectors 2023;16:206. [Google Scholar] | [Crossref]
- Nyangiwe N, Horak IG, Van der Mescht L, Matthee S. Range expansion of the economically important Asiatic blue tick,
Rhipicephalus microplus , in South Africa. J S Afr Vet Assoc 2017;88:a1414. [Google Scholar] | [Crossref] - Mpofu TJ, Nephawe KA, Mtileni B. Prevalence and resistance to gastrointestinal parasites in goats: A review. Vet World 2022;15(10):2442-2454. [Google Scholar] | [Crossref]
- Yawa M, Nyangiwe N, Jaja IF, Kadzere CT, Marufu MC. Communal cattle farmers'knowledge, attitudes and practices on ticks, tick control and acaricide resistance. Trop Anim Health Prod 2020;52:3005-3013. [Google Scholar] | [Crossref]
- Yawa M, Nyangiwe N, Jaja IF, Marufu MC, Kadzere CT. Acaricide resistance of
Rhipicephalus decoloratus ticks collected from communal grazing cattle in South Africa. J Adv Vet Anim Res 2022;9(1):33-41. [Google Scholar] | [Crossref] - Rodríguez-Vivas RI, Apanaskevich DA, Ojeda-Chi MM, Trinidad-Martínez I, Reyes-Novelo E, Esteve-Gassent MD. Ticks collected from humans, domestic animals and wildlife in Yucatan, Mexico. Vet Parasitol 2016;215:106-113. [Google Scholar] | [Crossref]
- Nyangiwe N, Yawa M, Qokweni L, Mthi S, Goni S, Mapholi NO. Species richness and adaptation of the invasive cattle tick
Rhipicephalus microplus in the Eastern Cape Province, South Africa. Ticks Tick Borne Dis 2023;14(3):102122. [Google Scholar] | [Crossref] - Nchu F, Nana P, Msalya G, Magano SR. Ethnoveterinary practices for control of ticks in Africa. In:EVM:Present and Future Concepts 2020:99-122. [Google Scholar] | [Crossref]
- Wesonga FD, Gachohi JM, Kitala PM, Gathuma JM, Njenga MJ. Seroprevalence of
Anaplasma marginale andBabesia bigemina infections and associated risk factors in Machakos County, Kenya. Trop Anim Health Prod 2017;49:265-272. [Google Scholar] | [Crossref] - Mutavi F, Heitkönig IMA, Wieland B, Aarts N, Van Paassen A. Tick treatment practices: Access to, knowledge about and on-farm use of acaricides in Laikipia, Kenya. Ticks Tick Borne Dis 2021;12(5):101757. [Google Scholar] | [Crossref]
- Abanda G, Josiane B. Tick-borne pathogens in African cattle: Novel molecular tools for diagnostics in epizootiology and the genetics of resistance. Doctoral dissertation, Universität Tübingen, Germany 2020. [Google Scholar] | [Crossref]
- Makwarela TG, Djikeng A, Masebe TM, Nyangiwe N, Nesengani LT, Mapholi NO. Vector abundance and abiotic factors influencing the distribution of ticks in six provinces of South Africa. Vet World 2024;17(8):1765-1776. [Google Scholar] | [Crossref]
- Davies S, Abdullah S, Helps C, Tasker S, Newbury H, Wall R. Prevalence of ticks and tick-borne pathogens (
Babesia andBorrelia spp.). in ticks infesting cats in Great Britain. Vet Parasitol 2017;244:129-135. [Google Scholar] | [Crossref] - Rajakaruna RS, Eremeeva ME. Eco-epidemiology of tick-borne pathogens: Role of tick vectors and host animal community composition. One Health Triad 2023:325-350. [Google Scholar] | [Crossref]
- Singh K, Kumar S, Sharma AK, Jacob SS, Verma MR, Singh NK. Economic impact of predominant ticks and tick-borne diseases on dairy production systems. Exp Parasitol 2022;243:108408. [Google Scholar] | [Crossref]
- Sajid MS, Rizwan HM, Khan MK, Qudoos A, Atif FA, Malik MA. Association of herd management with tick infestation in domestic goats. J Hellenic Vet Med Soc 2020;71(3):2283-2290. [Google Scholar] | [Crossref]
- Mtshali K, Khumalo ZT, Nakao R, Grab DJ, Sugimoto C, Thekisoe OMM. Molecular detection of zoonotic tick-borne pathogens in South Africa. J Vet Med Sci 2015;77(12):1573-1579. [Google Scholar] | [Crossref]
- Gray JS, Ogden NH. Ticks, human babesiosis and climate change. Pathogens 2021;10(11):1430. [Google Scholar] | [Crossref]
- Obaid MK, Islam N, Alouffi A, Khan AZ, da Silva Vaz I, Tanaka T. Acaricide resistance in ticks: Selection, diagnosis, mechanisms and mitigation. Front Cell Infect Microbiol 2022;12:941831. [Google Scholar] | [Crossref]
- Dzemo WD, Thekisoe O, Vudriko P. Development of acaricide resistance in cattle ticks: A systematic review and meta-analysis. Heliyon 2022;8(1):e08718. [Google Scholar] | [Crossref]
- Evans A, Madder M, Fourie J, Halos L, Kumsa B, Kimbita E. Acaricide resistance status of livestock ticks from East and West Africa. Int J Parasitol Drugs Drug Resist 2024;25:100541. [Google Scholar] | [Crossref]
- Achi LY, Boka M, Biguezoton A, Yao PK, Adakal H, Kande S. Resistance of
Rhipicephalus microplus to alphacypermethrin, deltamethrin and amitraz in Côte d'Ivoire. Int J Biol Chem 2022;16(3):910-922. [Google Scholar] | [Crossref] - Byaruhanga J, Vudriko P, Byamukama B, Odua F, Ssebunya Y, Njalira RK. Spatial distribution of tick acaricide resistance in Uganda. Open J Vet Med 2025;15(4):67-85. [Google Scholar] | [Crossref]
- Yawa M, Nyangiwe N, Muchenje V, Kadzere CT, Mpendulo TC, Marufu MC. Ecological preferences and seasonal dynamics of ticks on and off bovine hosts in the Eastern Cape Province, South Africa. Exp Appl Acarol 2018;74:317-328. [Google Scholar] | [Crossref]
- Nemaungwe TM, Van Dalen EM, Waniwa EO, Makaya PV, Chikowore G, Chidawanyika F. Biogeography of
Rhipicephalus appendiculatus under current and future climate scenarios of Zimbabwe. Exp Appl Acarol 2023;90(1):67-82. [Google Scholar] | [Crossref] - Patel MV, Kumar N, Solanki JB, Kalyani IH, Patel DC, Bhinsara DB. Relationship between flumethrin resistance and
Anaplasma marginale infection inRhipicephalus microplus . Trop Anim Health Prod 2024;56:113. [Google Scholar] | [Crossref] - Habeeba S, Mahmmod Y, Mohammed H, Amer H, Moustafa M, Sobhi A. In vitro detection of acaricide resistance in
Hyalomma species ticks. Vet Sci 2025;12(8):712. [Google Scholar] | [Crossref] - Sarwar MZ, Nomi ZA, Awais M, Shahbakht RM, Jamil M, Mussawar M. Effect of climate change on transmission of livestock diseases. Agrobiol Rec 2025;19:1-11. [Google Scholar] | [Crossref]
- Oli D, Gyawali B, Acharya S, Oshikoya S. Factors influencing learning attitude of farmers regarding adoption of farming technologies. Agric Technol 2025;10:2772-3755. [Google Scholar] | [Crossref]