


ABSTRACT
Background and Aim: Houseflies (
Materials and Methods: A cross-sectional surveillance study was conducted between June and November 2023 in Muang District, Phayao Province. A total of 350 houseflies were collected from meat, vegetable, and cooked-food markets using sticky traps.
Results: Overall, 106
Conclusion: Houseflies from food markets in Northern Thailand harbor virulent and MDR
Keywords: antimicrobial resistance,
INTRODUCTION
Antimicrobial resistance (AMR) has emerged as a critical global public health challenge and has received considerable attention from the World Health Organization (WHO) [1]. The increasing prevalence of pathogens exhibiting multidrug-resistant (MDR) phenotypes represents a major threat, as it substantially complicates the management of common infections and contributes to elevated morbidity, mortality, and healthcare costs [2, 3]. Owing to its far-reaching consequences for public health and global economic stability, the WHO has identified AMR as a strategic priority within the framework of the Sustainable Development Goals [4, 5]. Effective mitigation of AMR requires integrated approaches, including the establishment of robust surveillance systems, the promotion of targeted research, and the implementation of evidence-based public health interventions.
Houseflies are recognized mechanical vectors of a wide range of pathogens, including viruses, fungi, and bacteria such as
Despite the growing recognition of AMR as a major global health threat, surveillance efforts have largely focused on clinical, livestock, and food samples, with comparatively limited attention given to environmental vectors operating at the human–food interface. In particular, the role of houseflies as mechanical carriers of MDR
This study aimed to determine the prevalence of
MATERIALS AND METHODS
Ethical approval
This study did not require formal ethical approval from an Institutional Animal Ethics Committee or Institutional Review Board, as it exclusively involved the collection of houseflies (
Study period and location
This cross-sectional surveillance study was conducted from June to November 2023 in urban and peri-urban market settings in Muang District, Phayao Province, Northern Thailand.
Sample collection
Housefly samples were collected from local meat, vegetable, and cooked-food markets selected based on public accessibility, active food-handling activities, and their relevance as high-contact environments with potential environmental contamination and human exposure.
Sampling intensity was adjusted according to market size, with five traps deployed per sampling event in cooked-food markets (small), ten in vegetable markets (medium), and fifteen in meat markets (large). In total, 350 houseflies were collected using the sticky trap method, as previously described [11]. Sterile adhesive strips were placed within market areas, and captured houseflies were transferred individually into sterile containers using forceps. Samples were transported to the laboratory and euthanized at −20°C for 1–2 h. Morphological identification was performed using a stereomicroscope to confirm species identity as
Isolation of E. coli
A sterile loopful of enriched APW culture was streaked onto eosin methylene blue agar (HiMedia Laboratories Pvt. Ltd., Cat. No.M317-500G) and incubated at 37°C for 24 h. Presumptive
DNA extraction
Genomic DNA was extracted using the boiling method, as previously described [14]. A single colony of
Molecular confirmation and phylogenetic grouping
All isolates were molecularly confirmed as
Phylogenetic grouping was performed by multiplex PCR targeting the
Detection of virulence genes
Virulence genes associated with diarrheagenic
Antimicrobial susceptibility testing
Antimicrobial susceptibility was evaluated using the disk diffusion method in accordance with Clinical and Laboratory Standards Institute guidelines (CLSI M100-S23, 2018). Twelve antibiotics representing eight antimicrobial classes were tested: aminoglycosides (gentamicin, streptomycin), penicillins/β-lactams (penicillin G, ampicillin), carbapenems (imipenem, meropenem), tetracyclines (tetracycline, tigecycline), phenicols (chloramphenicol), fluoroquinolones (ciprofloxacin), macrolides (erythromycin), and sulfonamides (trimethoprim–sulfamethoxazole) (HiMedia Laboratories Pvt. Ltd., Cat. No. SD170, SD031, SD028, SD002, SD073, SD727, SD037, SD278, SD006, SD060, SD013, and SD010, respectively). Mueller–Hinton agar plates (HiMedia Laboratories Pvt. Ltd., Cat. No. M173) were inoculated with a 0.5 McFarland standardized suspension and incubated at 37°C for 18–24 h. Inhibition zones were measured in millimeters.
Detection of antimicrobial resistance genes
Phenotypically resistant
Statistical analysis
Descriptive data are presented as frequencies and percentages. Differences in categorical variables, including
RESULTS
Prevalence and phylogroup distribution of E. coli isolates
In this study, 106 (30.3%)
Table 1. Prevalence and occurrence rate of phylogenetic groups of
| Sampling area | Number of samples collected | A, n (%) | B1, n (%) | B2, n (%) | D, n (%) | |
|---|---|---|---|---|---|---|
| Meat market | 200 | 79 (39.5) | 42 (53.2) | 11 (13.9) | 7 (8.9) | 19 (24.1) |
| Vegetable market | 100 | 14 (14.0) | 7 (50.0) | 5 (35.7) | 0 (0.0) | 2 (14.3) |
| Cooked-food market | 50 | 13 (26.3) | 3 (23.1) | 4 (30.8) | 6 (46.2) | 0 (0.0) |
| Total | 350 | 106 (30.3) | 52 (49.1) | 20 (18.9) | 13 (12.3) | 21 (19.8) |
n = Number of isolates, % = Percentage within
Distribution of virulence genes in E. coli isolates
All
Table 2. Virulence gene profiles of
| Virulence profiles | Meat market (n = 79) | Vegetable market (n = 14) | Cooked-food market (n = 13) | Total (n = 106) | p-value |
|---|---|---|---|---|---|
| Negative | 24 (30.4) | 7 (50.0) | 1 (7.7) | 32 (30.2) | 0.0570 |
| Positive virulence | 55 (69.6) | 7 (50.0) | 12 (92.3) | 74 (69.8) | |
| One gene | |||||
| | 13 (16.5) | 0 (0.0) | 0 (0.0) | 13 (12.3) | |
| | 24 (30.4) | 1 (7.1) | 3 (23.1) | 28 (26.4) | |
| | 3 (3.8) | 0 (0.0) | 0 (0.0) | 3 (2.8) | |
| | 5 (6.3) | 0 (0.0) | 0 (0.0) | 5 (4.7) | |
| | 0 (0.0) | 0 (0.0) | 1 (7.7) | 1 (0.9) | |
| | 4 (5.1) | 4 (28.6) | 0 (0.0) | 8 (7.5) | |
| Two genes | |||||
| | 5 (6.3) | 1 (7.1) | 4 (30.8) | 10 (9.4) | |
| | 0 (0.0) | 0 (0.0) | 4 (30.8) | 4 (3.8) | |
| | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| | 0 (0.0) | 1 (7.1) | 0 (0.0) | 1 (0.9) |
n = Number of
Phylogenetic distribution of virulence-associated gene profiles
Virulence gene profiles varied across phylogenetic groups (Supplementary Table 2). Phylogroup A exhibited the greatest diversity of virulence-associated genes, with
Antibiotic resistance phenotype of E. coli isolates
Phenotypic antimicrobial resistance was widely observed among
Table 3. Antibiotic resistance profile of
| Drug classes | Antibiotics | Meat (n = 79) | Vegetable market (n = 14) | Cooked-food market (n = 13) | Total (n = 106) |
|---|---|---|---|---|---|
| Aminoglycosides | Gentamicin | 7 (8.9) | 0 (0.0) | 5 (38.5) | 12 (11.3) |
| Streptomycin | 12 (15.2) | 0 (0.0) | 0 (0.0) | 12 (11.3) | |
| Penicillin | Penicillin G | 79 (100.0) | 14 (100.0) | 13 (100.0) | 106 (100.0) |
| Ampicillin | 55 (69.9) | 7 (50.0) | 8 (61.5) | 70 (66.0) | |
| Carbapenem | Imipenem | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Meropenem | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | |
| Tetracycline | Tetracycline | 33 (41.8) | 0 (0.0) | 5 (38.5) | 38 (35.8) |
| Tigecycline | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) | |
| Phenicol | Chloramphenicol | 13 (16.5) | 0 (0.0) | 5 (38.5) | 18 (17.0) |
| Fluoroquinolone | Ciprofloxacin | 3 (3.8) | 0 (0.0) | 0 (0.0) | 3 (2.8) |
| Macrolide | Erythromycin | 79 (100.0) | 14 (100.0) | 13 (100.0) | 106 (100.0) |
| Sulfonamides | Trimethoprim/sulfamethoxazole | 10 (12.7) | 2 (14.3) | 0 (0.0) | 12 (11.3) |
Multidrug resistance was identified in 46.2% (49/106) of isolates (Table 4). The prevalence of MDR isolates was significantly higher in the meat market than in the vegetable and cooked-food markets (p = 0.0225). A total of 23 distinct AMR patterns were observed, with ampicillin–erythromycin–penicillin–tetracycline being the most frequent MDR profile (11.3%; 12/106). Only two isolates, both from the meat market, exhibited resistance to seven tested antibiotics.
Table 4. Antibiotic resistance patterns of
| Antibiotic-resistant patterns | Meat (n = 79) | Vegetable market (n = 14) | Cooked-food market (n = 13) | Total (n = 106) | p-value |
|---|---|---|---|---|---|
| MDR | 42 (53.2) | 2 (14.3) | 5 (38.5) | 49 (46.2) | 0.0225 |
| Two drugs | |||||
| E–P | 17 (21.5) | 7 (50.0) | 5 (38.5) | 29 (27.4) | |
| Three drugs | |||||
| AMP–E–P | 20 (25.3) | 5 (35.7) | 3 (23.1) | 28 (26.4) | |
| E–P–S | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| E–P–TE | 2 (2.5) | 0 (0.0) | 0 (0.0) | 2 (1.9) | |
| E–P–GEN | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| E–P–C | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| Four drugs | |||||
| AMP–E–P–TE | 12 (15.2) | 0 (0.0) | 0 (0.0) | 12 (11.3) | |
| AMP–E–P–SXT | 1 (1.3) | 2 (14.3) | 0 (0.0) | 3 (2.8) | |
| AMP–E–P–GEN | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| AMP–E–P–S | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| E–P–TE–C | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| Five drugs | |||||
| AMP–E–P–TE–SXT | 2 (2.5) | 0 (0.0) | 0 (0.0) | 2 (1.9) | |
| AMP–E–P–TE–S | 5 (6.3) | 0 (0.0) | 0 (0.0) | 5 (4.7) | |
| AMP–E–P–TE–C | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| AMP–E–P–S–SXT | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| E–P–TE–SXT | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| Six drugs | |||||
| AMP–E–P–TE–C–GEN | 4 (5.1) | 0 (0.0) | 5 (38.5) | 9 (8.5) | |
| AMP–E–P–C–SXT–CIP | 2 (2.5) | 0 (0.0) | 0 (0.0) | 2 (1.9) | |
| AMP–E–P–TE–S–C | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| AMP–E–P–TE–S–GEN | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| AMP–E–P–TE–S–SXT | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| Seven drugs | |||||
| AMP–E–P–TE–C–SXT–CIP | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| AMP–E–P–TE–C–SXT–S | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) |
MDR = Multidrug resistance, AMP = Ampicillin, E = Erythromycin, P = Penicillin G, TE = Tetracycline, S = Streptomycin, GEN = Gentamicin, C = Chloramphenicol, CIP = Ciprofloxacin, SXT = Trimethoprim/sulfamethoxazole, n = Number of
Distribution of MDR across phylogenetic groups and market types
Among the 106
Antibiotic resistance gene profiles of E. coli isolates
The distribution of AMR genes among resistant
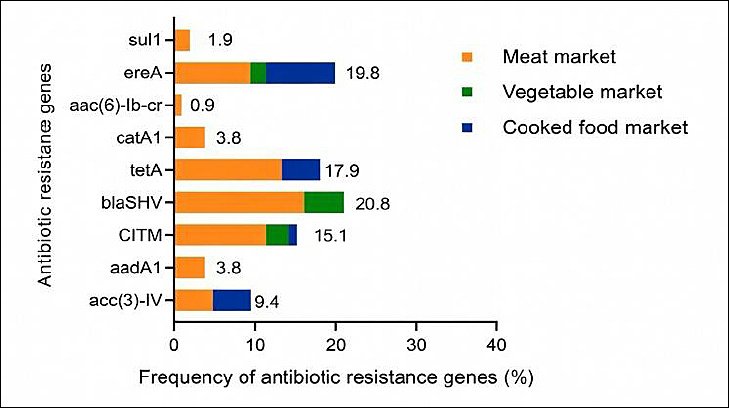
Figure 1. Resistance gene profiles were investigated among
Overall, 67 (63.2%) of the 106 isolates carried at least one AMR gene (Table 5). The most frequently detected single gene was
Table 5. Antibiotic resistance gene patterns of
| Antibiotic resistance gene expression patterns | Meat (n = 79) | Vegetable Market (n = 14) | Cooked-food market (n = 13) | Total (n = 106) | p-value |
|---|---|---|---|---|---|
| Negative | 34 (43.0) | 4 (25.6) | 1 (7.7) | 39 (36.8) | 0.0394 |
| Positive resistance genes | 45 (57.0) | 10 (97.4) | 12 (92.3) | 67 (63.2) | |
| One gene | |||||
| | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| | 2 (2.5) | 0 (0.0) | 0 (0.0) | 2 (1.9) | |
| | 8 (10.1) | 5 (35.7) | 0 (0.0) | 13 (12.3) | |
| | 3 (3.8) | 0 (0.0) | 0 (0.0) | 3 (2.8) | |
| | 5 (6.3) | 3 (21.4) | 1 (7.7) | 9 (8.5) | |
| | 4 (5.1) | 2 (14.3) | 6 (46.2) | 12 (11.3) | |
| | 6 (7.6) | 0 (0.0) | 0 (0.0) | 6 (5.7) | |
| Two genes | |||||
| | 3 (3.8) | 0 (0.0) | 2 (15.4) | 5 (4.7) | |
| | 4 (5.1) | 0 (0.0) | 0 (0.0) | 4 (3.8) | |
| | 2 (2.5) | 0 (0.0) | 0 (0.0) | 2 (1.9) | |
| Three genes | |||||
| | 2 (2.5) | 0 (0.0) | 0 (0.0) | 2 (1.9) | |
| | 2 (2.5) | 0 (0.0) | 3 (21.3) | 5 (4.7) | |
| | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) | |
| Four genes | |||||
| | 1 (1.3) | 0 (0.0) | 0 (0.0) | 1 (0.9) |
n = Number of
p-values were calculated using the chi-square test for comparisons of AMR gene-positive
DISCUSSION
Role of houseflies in pathogen transmission
Houseflies (
Prevalence of E. coli in market-associated houseflies
In this study,
Phylogenetic distribution of E. coli isolates
Phylogenetic analysis revealed that phylogroup A predominated among the
Virulence gene carriage and pathogenic potential
Virulence gene detection is essential for assessing the pathogenic potential of
The
Phylogenetic spread of diarrheagenic virulence markers
The detection of multiple DEC-associated virulence markers (
Antimicrobial resistance patterns in E. coli isolates
The detection of antibiotic-resistant
Multidrug resistance and phylogenetic associations
MDR was detected in 46.2% of isolates, with a significantly higher prevalence in meat markets than in other market types, reflecting environmental contamination and antimicrobial exposure [40]. Comparable MDR rates have been reported in urban environments [38], whereas lower rates were observed in dairy farm settings [41]. In contrast, higher MDR prevalence has been documented in hospital-adjacent environments [35]. The most frequent MDR pattern, ampicillin–erythromycin–penicillin–tetracycline, underscores the adaptability of
Antimicrobial resistance gene profiles and public health implications
AMR gene carriage was detected in 63.2% of isolates, highlighting the role of houseflies as environmental reservoirs of resistance determinants. The most prevalent gene was
CONCLUSION
This study demonstrated that
These findings highlight fresh-food markets, particularly meat markets, as critical interfaces for environmental dissemination of virulent and MDR
A key strength of this study lies in its integrated approach, combining prevalence estimation, phylogenetic grouping, virulence profiling, phenotypic resistance testing, and AMR gene detection within the same isolates. The comparison across different market types provides context-specific insights into exposure risk, while the focus on houseflies expands current understanding of non-clinical AMR reservoirs.
This cross-sectional study was confined to a single province, which may limit extrapolation to other regions. The absence of whole-genome sequencing restricted resolution of clonal relationships and mobile genetic elements. In addition, antimicrobial susceptibility was assessed using disc diffusion only, without MIC determination, and direct fly-to-food transmission and human exposure were not quantitatively evaluated.
Future studies should incorporate longitudinal sampling, whole-genome sequencing, and quantitative risk assessment to better elucidate transmission pathways of MDR
Overall, this study provides baseline evidence that
DATA AVAILABILITY
The supplementary data can be made available from the corresponding author upon request.
AUTHORS’ CONTRIBUTIONS
KK: Conceptualized and designed the study, collected samples, and performed the experiments. KK, SU, AS, and NS: Analyzed the data and drafted the manuscript. KK, SU, and NS: Reviewed and edited the manuscript. KK also secured funding for the study. OJ and CT: Data analysis. All authors have reviewed and approved the final manuscript of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
This research was supported by the University of Phayao and the Thailand Science Research and Innovation Fund (Fundamental Fund 2026, Grant No. 2260/2568 and FF65-RIM111).
REFERENCES
- Ajulo S, Awosile B. Global antimicrobial resistance and use surveillance system (GLASS 2022):investigating the relationship between antimicrobial resistance and antimicrobial consumption data across the participating countries. PLoS One 2024;19(2):e0297921. [Google Scholar] | [Crossref]
- Hariyanto H, Yahya CQ, Cucunawangsih C, Pertiwi CLP. Antimicrobial resistance and mortality. Afr J Infect Dis 2022;16(2):13-20. [Google Scholar] | [Crossref]
- Ahmed SK, Hussein S, Qurbani K, Ibrahim RH, Fareeq A, Kochr Mahmood KA. Antimicrobial resistance:impacts, challenges, and future prospects. J Med Sci Sur Public Health 2024;2:100081. [Google Scholar] | [Crossref]
- Uddin TM, Chakraborty AJ, Khusro A, Zidan BRM, Mitra S, Emran TB. Antibiotic resistance in microbes:history, mechanisms, therapeutic strategies and future prospects. J Infect Public Health 2021;14(12):1750-1766. [Google Scholar] | [Crossref]
- Bertagnolio S, Dobreva Z, Centner CM, Olaru ID, Donà D, Burzo S. WHO global research priorities for antimicrobial resistance in human health. Lancet Microbe 2024;5(11):100902. [Google Scholar] | [Crossref]
- Pakbin B, Brück WM, Rossen JWA. Virulence factors of enteric pathogenic
Escherichia coli :a review. Int J Mol Sci 2021;22:9922. [Google Scholar] | [Crossref] - Siriphap A, Suwancharoen C, Laenoi W, Kraivuttinun P, Suthienkul O, Prapasawat W. First study on virulence genes, antimicrobial resistance, and integrons in
Escherichia coli isolated from cage, free-range, and organic commercial eggs in Phayao Province, Thailand. Vet World 2022;15(9):2293-2301. [Google Scholar] | [Crossref] - Akter S, Sabuj AAM, Haque ZF, Kafi MA, Rahman MT, Saha S. Detection of antibiotic-resistant bacteria and their resistance genes from houseflies. Vet World 2020;13(2):266-274. [Google Scholar] | [Crossref]
- Park R, Dzialo MC, Spaepen S, Nsabimana D, Gielens K, Devriese H. Microbial communities of the house fly
Musca domestica vary with geographical location and habitat. Microbiome 2019;7:147. [Google Scholar] | [Crossref] - Sobur A, Haque ZF, Sabuj AA, Ievy S, Rahman AT, El Zowalaty ME. Molecular detection of multidrug and colistin-resistant
Escherichia coli isolated from house flies in various environmental settings. Future Microbiol 2019;14:847-858. [Google Scholar] | [Crossref] - Ranjbar R, Izadi M, Hafshejani TT, Khamesipour F. Molecular detection and antimicrobial resistance of
Klebsiella pneumoniae from house flies (Musca domestica ) in kitchens, farms, hospitals and slaughterhouses. J Infect Public Health 2016;9(4):499-505. [Google Scholar] | [Crossref] - Kababian M, Mozaffari E, Akbarzadeh K, Kordshouli RS, Saghafipour A, Shams S. Identification of bacteria contaminating
Musca domestica (Diptera:Muscidae) collected from animal husbandries. Shiraz E-Med J 2020;21(4):e92018. [Google Scholar] | [Crossref] - Hossain M, Rahman W, Ali M, Sultana T, Hossain K. Identification and antibiogram assay of
Escherichia coli isolated from chicken eggs. J Bio-Sci 2021;29:123-133. [Google Scholar] | [Crossref] - Dashti AA, Jadaon MM, Abdulsamad AM, Dashti HM. Heat treatment of bacteria:a simple method of DNA extraction for molecular techniques. Kuwait Med J 2009;41(2):117-122. [Google Scholar] | [Crossref]
- Heijnen L, Medema G. Quantitative detection of
E coli ,E. coli O157 and other shiga toxin producingE. coli in water samples using a culture method combined with real-time PCR. J Water Health 2006;4(4):487-498. [Google Scholar] | [Crossref] - Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of
Escherichia coli phylogenetic group. Appl Environ Microbiol 2000;66(10):4555-4558. [Google Scholar] | [Crossref] - Performance standards for antimicrobial susceptibility testing:twenty-third informational supplement. Wayne, PA: Clinical and Laboratory Standards Institute; 2018. [Google Scholar]
- Jain P, Bepari AK, Sen PK, Rafe T, Imtiaz R, Reza HM. High prevalence of multiple antibiotic resistance in clinical
E coli isolates from Bangladesh and prediction of molecular resistance determinants using WGS of an XDR isolate. Sci Rep 2021;11:22859. [Google Scholar] | [Crossref] - Van TTH, Chin J, Chapman T, Tran LT, Coloe PJ. Safety of raw meat and shellfish in Vietnam:an analysis of
Escherichia coli isolations for antibiotic resistance and virulence genes. Int J Food Microbiol 2008;124(3):217-223. [Google Scholar] | [Crossref] - Park CH, Robicsek A, Jacoby GA, Sahm DC. Prevalence in the United States of
aac(6')-Ib-cr encoding a ciprofloxacin-modifying enzyme. Antimicrob Agents Chemother 2006;50(11):3953-3955. [Google Scholar] | [Crossref] - Smallegange RC, Den Otter CJ, Takken W, Knols BGJ. Houseflies, annoying and dangerous. Wageningen: Wageningen Academic Publishers; 2007. p. 281-292. [Google Scholar]
- Nazari M, Mehrabi T, Hosseini SM, Alikhani MY. Bacterial contamination of adult house flies (
Musca domestica ) and sensitivity of these bacteria to various antibiotics, captured from Hamadan city, Iran. J Clin Diagn Res 2017;11((4)):DC04. [Google Scholar] | [Crossref] - Chandrakar C, Shakya S, Patyal A, Jain A, Ali SL, Mishra OP. ERIC-PCR-based molecular typing of multidrug-resistant
Escherichia coli isolated from houseflies (Musca domestica ) in the environment of milk and meat shops. Lett Appl Microbiol 2022;75(6):1549-1558. [Google Scholar] | [Crossref] - Zuhora FT, Hosen MA, Rumi NA, Rahman MS, Alam J, Hasan MH. Molecular characterization of multidrug-resistant bacteria isolated from the external and internal parts of the housefly. J Adv Biotechnol Exp Ther 2023;6(3):597-609. [Google Scholar] | [Crossref]
- Liu Y, Yang Y, Zhao F, Fan X, Zhong W, Qiao D. Multi-drug resistant Gram-negative enteric bacteria isolated from flies at Chengdu Airport, China. Southeast Asian J Trop Med Public Health 2013;44(6):988-996. [Google Scholar] | [Crossref]
- Mersha G, Asrat D, Zewde BM, Kyule M. Occurrence of
Escherichia coli O157:H7 in feces, skin and carcasses from sheep and goats in Ethiopia. Lett Appl Microbiol 2010;50(1):71-76. [Google Scholar] | [Crossref] - Messele YE, Abdi RD, Yalew ST, Tegegne DT, Emeru BA, Weird GM. Molecular determination of antimicrobial resistance in
Escherichia coli isolated from raw meat in Addis Ababa and Bishoftu, Ethiopia. Ann Clin Microbiol Antimicrob 2017;16:55. [Google Scholar] | [Crossref] - Stoppe NC, Silva JS, Carlos C, Sato MIZ, Saraiva AM, Ottoboni LMM. Worldwide phylogenetic group patterns of
Escherichia coli from commensal human and wastewater treatment plant isolates. Front Microbiol 2017;8:2512. [Google Scholar] | [Crossref] - Mosquito S, Pons MJ, Riveros M, Ruiz J, Ochoa TJ. Diarrheagenic
Escherichia coli phylogroups are associated with antibiotic resistance and duration of diarrheal episode. Sci World J 2015;2015:610403. [Google Scholar] | [Crossref] - Robert E, Grippa M, Nikiema DE, Kergoat L, Koudougou H, Auda Y. Environmental determinants of
E coli , link with the diarrheal diseases, and indication of vulnerability criteria in tropical West Africa (Kapore, Burkina Faso). PLoS Negl Trop Dis 2021;15(8):e0009634. [Google Scholar] | [Crossref] - Sidhu JP, Ahmed W, Hodgers L, Toze S. Occurrence of virulence genes associated with diarrheagenic pathotypes in
Escherichia coli isolates from surface water. Appl Environ Microbiol 2013;79(1):328-335. [Google Scholar] | [Crossref] - Nataro JP, Kaper JB. Diarrheagenic
Escherichia coli . Clin Microbiol Rev 1998;11(1):142-201. [Google Scholar] | [Crossref] - Sobur MA, Sabuj AAM, Sarker R, Rahman AMMT, Kabir SML, Rahman MT. Antibiotic-resistant
Escherichia coli andSalmonella spp. associated with dairy cattle and farm environment having public health significance. Vet World 2019;12(7):984-993. [Google Scholar] | [Crossref] - Fujioka M, Otomo Y, Ahsan CR. A novel single-step multiplex polymerase chain reaction assay for the detection of diarrheagenic
Escherichia coli . J Microbiol Methods 2013;92(3):289-292. [Google Scholar] | [Crossref] - Afset JE, Anderssen E, Bruant G, Harel J, Wieler L, Bergh K. Phylogenetic backgrounds and virulence profiles of atypical enteropathogenic
Escherichia coli strains from a case-control study using multilocus sequence typing and DNA microarray analysis. J Clin Microbiol 2008;46(7):2280-2290. [Google Scholar] | [Crossref] - Kaye KS, Gupta V, Mulgirigama A, Joshi AV, Scangarella-Oman NE, Yu K. Antimicrobial resistance trends in urine
Escherichia coli isolates from adult and adolescent females in the United States from 2011 to 2019:rising ESBL strains and impact on patient management. Clin Infect Dis 2021;73(11):1992-1999. [Google Scholar] | [Crossref] - Siriphap A, Kitti T, Khuekankaew A, Boonlao C, Thephinlap C, Thepmalee C. High prevalence of extended-spectrum beta-lactamase-producing
Escherichia coli andKlebsiella pneumoniae isolates:a 5-year retrospective study at a tertiary hospital in Northern Thailand. Front Cell Infect Microbiol 2022;12:955774. [Google Scholar] | [Crossref] - Zuhora FT, Hosen MB, Rumi NA, Rahman MT, Alam J, Hasan MR. Molecular characterization of multidrug-resistant bacteria isolated from the external and internal parts of the housefly. J Adv Biotechnol Exp Ther 2023;6(3):597. [Google Scholar] | [Crossref]
- Odetoyin B, Adeola B, Olaniran O. Frequency and antimicrobial resistance patterns of bacterial species isolated from the body surface of the housefly (
Musca domestica ) in Akure, Ondo State, Nigeria. J Arthropod Borne Dis 2020;14(1):88-96. [Google Scholar] | [Crossref] - Songe MM, Hang'ombe BM, Knight-Jones TJ, Grace D. Antimicrobial-resistant enteropathogenic
Escherichia coli andSalmonella spp. in houseflies infesting fish in food markets in Zambia. Int J Environ Res Public Health 2017;14(1):21. [Google Scholar] | [Crossref] - Dos Santos Alves TS, Lara GHB, Maluta RP, Ribeiro MG, Leite DS. Carrier flies of multidrug-resistant
Escherichia coli as potential dissemination agent in dairy farm environment. Sci Total Environ 2018;633:1345-1351. [Google Scholar] | [Crossref] - Mathers AJ, Peirano G, Pitout JD. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant
Enterobacteriaceae . Clin Microbiol Rev 2015;28(3):565-591. [Google Scholar] | [Crossref] - Skurnik D, Lacheeb S, Bernede C, le Menac'h A, Elbaz S, Mohler J. Integrons and antibiotic resistance in phylogenetic group B2
Escherichia coli . Microb Drug Resist 2009;15(3):173-178. [Google Scholar] | [Crossref] - Petty NK, Ben Zakour NL, Stanton-Cook M, Skippington E, Totsika M, Forde BM. Global dissemination of a multidrug resistant
Escherichia coli clone. Proc Natl Acad Sci U S A 2014;111(15):5694-5699. [Google Scholar] | [Crossref] - Bertelloni F, Bresciani F, Cagnoli G, Scotti B, Lazzerini L, Marcucci M. House flies (
Musca domestica ) from swine and poultry farms carrying antimicrobial resistantEnterobacteriaceae andSalmonella . Vet Sci 2023;10(2):118. [Google Scholar] | [Crossref]