


ABSTRACT
Background and Aim: After more than three decades of freedom from foot-and-mouth disease (FMD), Indonesia experienced widespread outbreaks in 2022–2023, raising major concerns regarding viral evolution and vaccine effectiveness. Foot-and-mouth disease virus (FMDV) serotype O remains the predominant circulating serotype in the region. However, the immunological and structural consequences of recent genetic variation have not been comprehensively evaluated. This study aimed to integrate molecular, immunoinformatic, and structural analyses to characterize FMDV serotype O isolates from West Java and South Sumatra and to assess their implications for antigenicity, immune recognition, and vaccine matching.
Materials and Methods: Clinical epithelial samples were collected from naturally infected cattle during outbreaks in West Java and South Sumatra. Viral RNA was extracted, and the capsid genes
Results: Sequence comparison revealed reduced identity of
Conclusion: This integrated molecular–immunoinformatic–structural analysis demonstrates that newly circulating Indonesian FMDV serotype O isolates exhibit genetic, antigenic, and structural divergence that may reduce current vaccine matching. Continuous molecular surveillance and regionally adapted vaccine design are therefore essential to maintain effective FMD control in Indonesia.
Keywords: foot-and-mouth disease virus, immunoinformatics, Indonesia, molecular docking, phylogenetic analysis, serotype O, TLR7 interaction,
INTRODUCTION
Foot-and-mouth disease (FMD) is a highly contagious viral disease affecting cloven-hoofed livestock, including cattle, goats, sheep, and pigs, and it also infects wild ungulates, particularly species at risk of extinction [1]. Caused by an aphthovirus, FMD spreads rapidly through direct contact with infected animals or via contaminated fomites. Clinically, the disease is characterized by fever, vesicular lesions in the oral cavity and feet, lameness, and reduced milk yield, leading to substantial economic losses for livestock producers. Vaccination programs combined with strict biosecurity measures are therefore essential for effective disease prevention and control. Although FMD has been eradicated in several regions, including Australia, Canada, Greenland, Mexico, and New Zealand, it remains a major transboundary animal disease threat in many parts of Africa, Asia, and South America.
Foot-and-mouth disease virus (FMDV) is a positive-sense single-stranded RNA virus with a genome of approximately 8400 nucleotides. Viral particles are spherical, with diameters ranging from 25 to 30 nm, and consist of an RNA genome enclosed within a protein capsid. The capsid is composed of 60 capsomeres, each containing four structural polypeptides: VP1, VP2, VP3, and VP4. The proteins VP1, VP2, and VP3 are exposed on the viral surface, whereas
In 2022, FMD outbreaks continued to affect several Asian countries, including Cambodia, China, peninsular Malaysia, Indonesia, Thailand, and Vietnam, whereas no new outbreaks were reported in Lao PDR, Mongolia, or Myanmar. The importation of cattle and meat products markedly increases the risk of FMD transmission, as the virus can disseminate rapidly through livestock movement between regions. After more than 30 years of disease freedom, Indonesia experienced a major re-emergence of FMD in 2022. The outbreak initially reappeared in Gresik, East Java, and subsequently spread swiftly across multiple provinces, including West Java, Bali, West Kalimantan, Central Kalimantan, East Kalimantan, and West Nusa Tenggara, ultimately affecting almost all provinces on Sumatra.
Phylogenetic analysis reported in a previous study [6] indicated that the virus responsible for the 2022 outbreak originated from Gresik and Banjarnegara in East Java Province. Clinical samples analyzed in the present study were obtained from FMD outbreaks in Indonesia during 2022–2023. Indonesia is now considered endemic for FMD, and the introduction of novel viral lineages into Southeast Asia has been increasingly reported [6].
Despite extensive molecular surveillance of FMDV in Southeast Asia, most studies conducted after the 2022 re-emergence in Indonesia have primarily focused on partial genomic characterization and lineage assignment based on
The present study aimed to comprehensively characterize FMDV serotype O isolates collected during the 2022–2023 outbreaks in Indonesia using an integrated molecular, immunoinformatic, and structural framework. Specifically, this study sought to (i) determine the genetic diversity and phylogenetic relationships of circulating isolates based on
MATERIALS AND METHODS
Ethical approval
All animal procedures were conducted in accordance with Indonesia’s national guidelines for animal welfare and biosafety for transboundary animal diseases. The study protocol, including field sampling, animal handling, and laboratory processing of infectious materials, was reviewed and approved by the Ethical Clearance Committee of the National Research and Innovation Agency (BRIN), Indonesia (approval no. 065/KE.02/SK/10/2022, October 6, 2022). Only naturally infected animals identified during the FMD outbreak were included; no experimental infection, disease induction, or invasive procedures were performed. Oropharyngeal epithelium and lesion scrapings were collected by licensed veterinarians and trained animal health officers following World Organisation for Animal Health (WOAH) guidelines for safe FMD sampling. Animals were handled using minimal-restraint techniques without sedation to avoid pain or distress. Farmers or animal owners provided prior informed consent, and all activities were supervised by the District Livestock Services in West Java and South Sumatra. Sample collection was conducted using appropriate personal protective equipment, and transport complied with national regulations for infectious substances. Laboratory work, including RNA extraction, cDNA synthesis, polymerase chain reaction (PCR) amplification, and sample storage, was performed in a BSL-2 facility with Class II biological safety cabinets. Waste materials were decontaminated by autoclaving or chemical disinfection. The study adhered to Animal Research: Reporting of
Study period and location
Samples were obtained from suspected FMD cases based on clinical signs (hypersalivation and vesicular lesions of the oral cavity and feet) in West Java and South Sumatra, Indonesia, during 2022–2023 (Figure 1). Sampling was conducted on smallholder cattle farms raising local breeds (
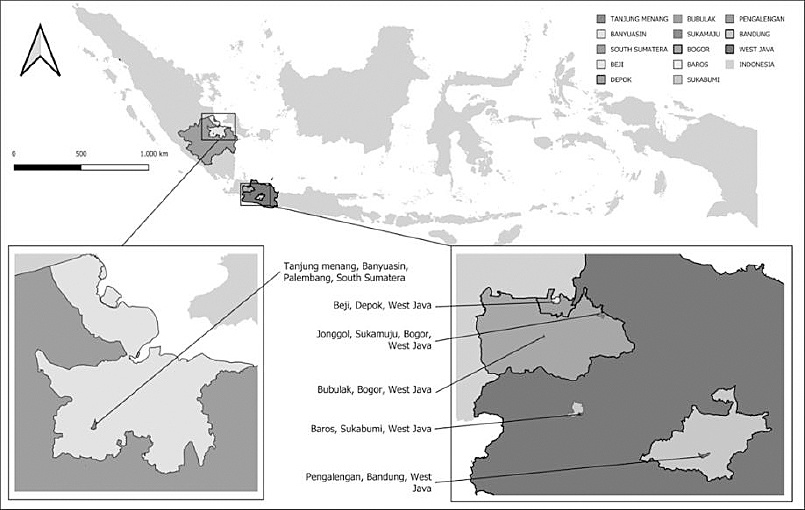
Figure 1. The geographical location of the sample collection areas in West Java and South Sumatra.
Sample collection and transportation
The methodology used in this study is presented in Figure 2. Animals with clinical scores of 2 and 3 (moderate and severe, respectively) presenting vesicles and ulceration of the oral cavity and interdigital spaces of the feet, hypersalivation, and lameness were selected. A total of 28 clinical samples (oral and foot epithelium) were collected and preserved in RNA Shield (Zymo Research, Irvine, CA, USA) (catalog no. R1100-50). The samples were transported in insulated containers with ice packs at approximately 4°C, with temperature validation using temperature logs, over 48–72 h. RNA Shield (Zymo Research) was used to protect nucleic acids at ambient temperature for up to 7 days. All samples were stored at −20°C in the laboratory for further analysis.
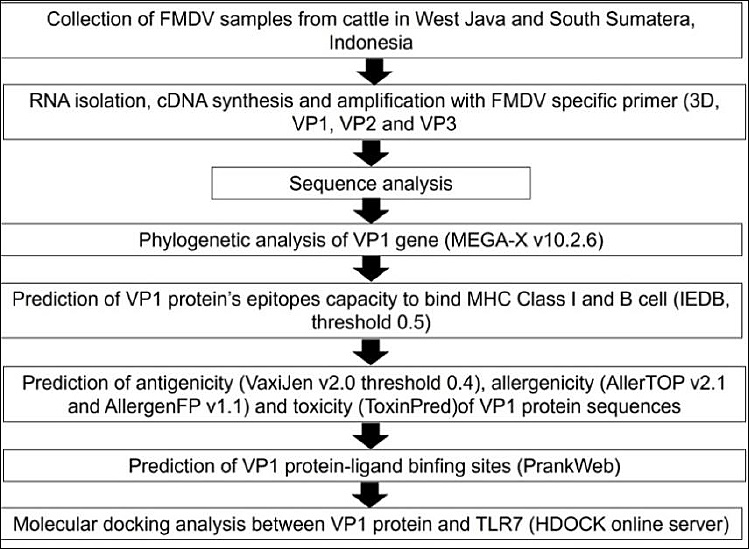
Figure 2. The methodology schematic used in this study.
RNA extraction and cDNA synthesis
Total RNA extraction was performed according to the manufacturer’s instructions using the Quick-RNA Miniprep Kit (Zymo Research) (catalog no. R1054), with a total RNA elution volume of 30 µL. RNA quality and concentration were evaluated using a NanoDrop spectrophotometer, and RNA with an A260/280 ratio of 1.8–2.0 was selected for further processing. cDNA synthesis was performed using the RevertAid cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA, USA) (catalog no. K1622) to generate complementary DNA. Negative controls (RNase/DNase-free water) were included to exclude contamination. The reaction setup involved incubation at 65°C for 5 min, followed by cDNA synthesis at 25°C for 5 min, 42°C for 60 min, and enzyme inactivation at 70°C for 5 min.
PCR amplification and capsid gene analysis
The cDNA was amplified by conventional PCR using DreamTaq PCR Master Mix (Thermo Fisher Scientific) (catalog no. K1081) with 3D primers for FMD diagnosis [7]. PCR conditions included initial denaturation at 95°C for 5 min, followed by 35 cycles of denaturation at 95°C for 10 s, annealing at 60°C for 20 s, and extension at 72°C for 30 s, with a final extension at 72°C for 5 min. The capsid genes in positive samples were analyzed using primers designed based on the FMDV serotype O reference sequence (MN095357.1) (Table 1). Gene-specific annealing temperatures were applied for
Table 1. Primers used for foot-and-mouth disease virus capsid diagnosis and analysis.
| Gene | Primer sequence (5′–3′) |
|---|---|
| 3D | Forward: ACTGGGTTTTACAAACCTGTGA |
| Reverse: GCGAGTCCTGCCACGGA | |
| VP1 | Forward: CTTCTCCGACGTTAGGTC |
| Reverse: GTCACGTGCTTTGAGCTG | |
| VP2 | Forward: CTTCTCCGACGTTAGGTC |
| Reverse: GTCACGTGCTTTGAGCTG | |
| VP3 | Forward: CTTCTCCGACGTTAGGTC |
| Reverse: GTCACGTGCTTTGAGCTG |
Sequence quality assessment and data submission
Sequence data were analyzed using MEGA-X v10.2.6. Low-quality regions were trimmed based on chromatogram inspection, Phred quality scores
Phylogenetic analysis of VP1
Aligned
Epitope prediction for MHC class I and B cells
Translated
Antigenicity, allergenicity, and toxicity assessment
Antigenicity was evaluated using VaxiJen v2.0 [11]. Allergenicity was assessed using AllerTOP v2.1 and AllergenFP v1.1 [12, 13]. Toxicity prediction was conducted using ToxinPred [14]. Predicted epitopes were benchmarked against experimentally validated epitopes.
Prediction of VP1 ligand-binding sites
Ligand-binding site prediction and 3D visualization were performed using PrankWeb [15].
Molecular docking between VP1 and TLR7
Molecular docking between
RESULTS
Detection and sequencing of FMDV serotype O capsid genes
Local FMDV serotype O strains were obtained from naturally infected cattle in South Sumatra and West Java during the 2022–2023 outbreak period (Table 2). PCR amplification was performed using serotype O–specific primers targeting the capsid gene coding regions
Table 2. Foot-and-mouth disease virus diagnosis and
| Sample code | City/Province |
|
|
|
|
|---|---|---|---|---|---|
| Bandung 1 | Bandung, West Java, Indonesia | + | + | + | + |
| Bandung 2 | Bandung, West Java, Indonesia | − | − | − | − |
| Bandung 3 | Bandung, West Java, Indonesia | + | + | + | + |
| Bandung 4 | Bandung, West Java, Indonesia | + | + | + | + |
| Bandung 5 | Bandung, West Java, Indonesia | + | + | + | + |
| Bandung 6 | Bandung, West Java, Indonesia | − | − | − | − |
| Bandung 7 | Bandung, West Java, Indonesia | − | − | − | − |
| Bandung 8 | Bandung, West Java, Indonesia | + | + | + | + |
| Bandung 9 | Bandung, West Java, Indonesia | + | + | + | + |
| Palembang 1 | Palembang, South Sumatra, Indonesia | + | + | + | + |
| Palembang 2 | Palembang, South Sumatra, Indonesia | + | + | + | + |
| Palembang 3 | Palembang, South Sumatra, Indonesia | + | + | + | + |
| Palembang 4 | Palembang, South Sumatra, Indonesia | + | + | + | + |
| Palembang 5 | Palembang, South Sumatra, Indonesia | − | − | − | − |
| Palembang 6 | Palembang, South Sumatra, Indonesia | + | + | + | + |
| Palembang 7 | Palembang, South Sumatra, Indonesia | + | + | + | + |
| Palembang 8 | Palembang, South Sumatra, Indonesia | + | + | + | + |
| Palembang 9 | Palembang, South Sumatra, Indonesia | + | + | + | + |
| Bogor 1 | Bogor, West Java, Indonesia | + | + | + | + |
| Bogor 2 | Bogor, West Java, Indonesia | − | − | − | − |
| Bogor 3 | Bogor, West Java, Indonesia | − | − | − | − |
| Bogor 4 | Bogor, West Java, Indonesia | − | − | − | − |
| Bogor 5 | Bogor, West Java, Indonesia | − | − | − | − |
| Bogor 6 | Bogor, West Java, Indonesia | − | − | − | − |
| Depok 1 | Depok, West Java, Indonesia | + | + | + | + |
| Depok 2 | Depok, West Java, Indonesia | + | + | + | + |
| Sukabumi 1 | Sukabumi, West Java, Indonesia | + | + | + | + |
| Sukabumi 2 | Sukabumi, West Java, Indonesia | + | + | + | + |
+ detected, − undetected
Table 3. Comparison of gene sequence similarity between collected samples and recent FMDV O isolate ISA/1/2022.
| Sample |
|
|
|
|---|---|---|---|
| Bandung 1 | 99.68% | 99.85% | 98.64% |
| Bandung 3 | 99.68% | 99.85% | 98.64% |
| Bandung 4 | 99.68% | 99.85% | 98.92% |
| Bandung 5 | 99.68% | 99.85% | 98.92% |
| Bandung 8 | 99.68% | 99.85% | 98.92% |
| Bandung 9 | 99.21% | 99.38% | 98.92% |
| Palembang 1 | 99.05% | 99.85% | 98.48% |
| Palembang 2 | 99.05% | 99.85% | 98.48% |
| Palembang 3 | 99.05% | 99.85% | 98.48% |
| Palembang 4 | 99.05% | 99.22% | 98.93% |
| Palembang 6 | 99.05% | 99.85% | 98.48% |
| Palembang 7 | 99.05% | 99.19% | 98.93% |
| Palembang 8 | 98.58% | 99.85% | 98.48% |
| Palembang 9 | 99.05% | 99.85% | 98.48% |
| Bogor 1 | 99.68% | 99.69% | 99.53% |
| Depok 1 | 99.68% | 99.69% | 99.53% |
| Depok 2 | 98.26% | 99.03% | 98.59% |
| Sukabumi 1 | 98.42% | 98.77% | 98.73% |
| Sukabumi 2 | 98.42% | 98.77% | 98.73% |
FMDV = Foot-and-mouth disease virus, ISA = Indonesian isolate.
Phylogenetic analysis of VP1
Phylogenetic reconstruction based on nucleotide substitution models classified the recent FMDV serotype O isolates into three principal clusters. Cluster I comprised Depok 2, Sukabumi 1, Sukabumi 2, and reference FMDV serotype O strains from the database (O/ME-SA/Ind-2001d, Myanmar, India, Oman, Kuwait, Iran, and Bhutan). Cluster II included Bandung 9 and Palembang 1, 2, 3, 4, 6, 7, and 8. Cluster III consisted of Bogor 1, Bandung 1, 3, 4, 5, 8, and Depok 1, which showed close genetic relatedness to Indonesian reference isolates ISA/Gresik/2022 and ISA/Banjarnegara/2022. In contrast, phylogenetic analysis based on amino acid substitutions resolved only two main clusters: Cluster I included Depok 2, Sukabumi 1, Sukabumi 2, and related database isolates, whereas Cluster II comprised the remaining samples together with ISA/Gresik/2022 and ISA/Banjarnegara/2022 (Figure 3).

Figure 3. Phylogenetic tree using MEGA-X v10.2.6, the maximum likelihood method based on full
Comparative analysis of
Table 4. Nucleic acid variation in the
| Position of nucleic acid | ISA/Indonesia 2022 | Bandung 1, 3, 4 | Bandung 5 | Bandung 8 | Bandung 9 | Sukabumi 1, 2 | Bogor 1 | Depok 2 | Depok 1 | Palembang 1, 2, 3, 6, 9 | Palembang 4, 7 | Palembang 8 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10 | G | A | A | A | A | A | A | A | A | A | A | A |
| 37 | G | G | G | G | G | A | G | A | G | G | G | G |
| 84 | G | G | G | G | G | G | G | G | G | G | A | G |
| 102 | T | T | T | T | T | C | T | C | T | T | T | T |
| 103 | A | A | A | A | G | A | A | A | A | A | G | A |
| 171 | C | C | C | C | C | T | C | T | C | C | C | C |
| 219 | C | C | C | C | C | T | C | T | C | C | C | C |
| 225 | C | C | C | C | C | T | C | T | C | C | C | C |
| 386 | C | C | C | C | T | T | T | T | C | T | T | T |
| 408 | C | C | C | C | C | C | C | T | C | C | C | C |
| 451 | T | C | C | C | C | T | T | T | T | C | C | C |
| 459 | G | G | G | C | G | G | G | G | G | G | G | T |
| 462 | G | G | G | G | G | G | G | G | G | G | G | T |
| 465 | A | A | A | A | A | A | A | A | A | G | A | G |
| 515 | G | G | G | G | G | A | G | A | G | G | G | G |
| 526 | T | T | T | T | T | T | T | T | T | C | T | C |
| 582 | T | T | C | T | T | T | T | T | T | T | T | T |
| 589 | G | G | G | G | G | G | G | G | G | G | G | A |
| 615 | C | C | C | C | C | T | C | T | C | C | C | C |
| 627 | C | C | C | C | C | T | C | T | C | C | C | C |
ISA = Indonesian isolate.
Table 5. Amino acid variation in VP1 protein between collected samples and ISA/Indonesia 2022 isolate.
| Amino acid position | ISA/Indonesia 2022 | Bandung 1, 3, 4, 5, 8, 9 | Sukabumi 1, 2 | Bogor 1 | Depok 2 | Depok 1 | Palembang 1, 2, 3, 4, 6, 7, 9 | Palembang 8 |
|---|---|---|---|---|---|---|---|---|
| 4 | A | T | T | T | T | T | T | T |
| 13 | A | A | T | A | T | A | A | A |
| 129 | A | A | V | V | V | A | V | V |
| 172 | R | R | Q | R | Q | R | R | R |
| 197 | E | E | E | E | E | E | E | K |
ISA = Indonesian isolate
Epitope prediction and immunological profiling of VP1
Based on MHC class I–binding predictions, the VP1 protein yielded eight peptides capable of binding T-cell epitopes associated with BoLA-1, BoLA-2, BoLA-3, BoLA-4, BoLA-5, BoLA-6, BoLA-T5, and BoLA-T7 alleles. Compared with Ind/2022, ISA/Banjarnegara/Gresik, and other FMDV serotype O isolates, samples from Bandung 1, 3, 4, 5, 8, 9, and Palembang 8 exhibited distinct T-cell epitope binding profiles. Samples Depok 2 and Sukabumi 1 and 2 differed from Ind/2022 and ISA/Banjarnegara/Gresik only at amino acid positions 4 (alanine) and 13 (tyrosine), localized within residues 2–15 of
Table 6. Prediction of
| BoLA allele (position) | Peptide (aa position) | Peptide sequence | Databases and code samples | Antigenicity score | Antigenicity | Allergenicity | Toxicity | Percentile rank |
|---|---|---|---|---|---|---|---|---|
| BoLA-1:00901 (69–82) | 69–82 | ATYYFADLEVAVKH | Strain O/ME-SA/Ind, MYA/13/2017, PD287/2015, PD73/2017, IRN/72/2009, BHU/3/2009, O/OMN/7/2001, O/KUW/3/97, INDIA 2013, ISA/GRESIK/2022, ISA/BANJARNEGARA/2022, INDONESIA 2022, Bandung 1, 3, 4, 5, 8, 9, Depok 1, 2, Bogor 1, Sukabumi 1, 2, Palembang 1, 2, 3, 4, 6, 7, 8, 9 | 0.9473 | Antigen | Non-allergen | Non-toxin | 2.6 |
| ATYYFAELEVAVKH | PD118/2017 (India) | 0.7559 | Antigen | Allergen | Non-toxin | 2.5 | ||
| BoLA-2:00501 (38–51) | 38–51 | RFVKVTPKDQINVL | Strain O/ME-SA/Ind, MYA/13/2017, PD287/2015, PD118/2017, PD73/2017, IRN/72/2009, BHU/3/2009, O/OMN/7/2001, O/KUW/3/97, INDIA 2013, ISA/GRESIK/2022, ISA/BANJARNEGARA/2022, INDONESIA 2022, Bandung 1, 3, 4, 5, 8, 9, Depok 1, 2, Bogor 1, Sukabumi 1, 2, Palembang 1, 2, 3, 4, 6, 7, 8, 9 | 0.8011 | Antigen | Non-allergen | Non-toxin | 3.9 |
| BoLA-3:00101 (38–51) | 38–51 | RFVKVTPKDQINVL | MYA/13/2017, PD287/2015, PD118/2017, PD73/2017, IRN/72/2009, BHU/3/2009, O/OMN/7/2001, O/KUW/3/97, INDIA 2013, ISA/GRESIK/2022, ISA/BANJARNEGARA/2022, INDONESIA 2022, Bandung 1, 3, 4, 5, 8, 9, Depok 1, 2, Bogor 1, Sukabumi 1, 2, Palembang 1, 2, 3, 4, 6, 7, 8, 9 | 0.8011 | Antigen | Non-allergen | Non-toxin | 13 |
| BoLA-4:02401 (75–88) | 75–88 | DLEVAVKHEGNLTW | Strain O/ME-SA/Ind, MYA/13/2017, PD287/2015, PD73/2017, IRN/72/2009, BHU/3/2009, O/OMN/7/2001, O/KUW/3/97, ISA/GRESIK/2022, ISA/BANJARNEGARA/2022, INDONESIA 2022, Bandung 1, 3, 4, 5, 8, 9, Depok 1, 2, Bogor 1, Sukabumi 1, 2, Palembang 1, 2, 3, 4, 6, 7, 8, 9 | 1.4045 | Antigen | Allergen | Non-toxin | 4.9 |
| ELEVAVKHEGNLTW | PD118/2017 (India) | 0.7559 | Antigen | Allergen | Non-toxin | 3.3 | ||
| DLEVAVKHKGNLTW | INDIA 2013 | 1.6457 | Antigen | Allergen | Non-toxin | 5.4 | ||
| BoLA-5:00301 (178–191) | 178–191 | RMKRAETYCPRPLL | Strain O/ME-SA/Ind, MYA/13/2017, PD287/2015, PD118/2017, PD73/2017, IRN/72/2009, BHU/3/2009, O/OMN/7/2001, O/KUW/3/97, INDIA 2013, ISA/GRESIK/2022, ISA/BANJARNEGARA/2022, INDONESIA 2022, Bandung 1, 3, 4, 5, 8, 9, Depok 1, 2, Bogor 1, Sukabumi 1, 2, Palembang 1, 2, 3, 4, 6, 7, 8, 9 | 0.6731 | Antigen | Allergen | Non-toxin | 8.9 |
| BoLA-6:01301 (178–191) | 178–191 | RMKRAETYCPRPLL | Same as above | 0.6731 | Antigen | Allergen | Non-toxin | 0.61 |
| BoLA-T5 (69–82) | 69–82 | ATYYFADLEVAVKH | Strain O/ME-SA/Ind, MYA/13/2017, PD287/2015, PD73/2017, IRN/72/2009, BHU/3/2009, O/OMN/7/2001, O/KUW/3/97, INDIA 2013, ISA/GRESIK/2022, ISA/BANJARNEGARA/2022, INDONESIA 2022, Bandung 1, 3, 4, 5, 8, 9, Depok 2, Sukabumi 1, 2, Palembang 8 | 0.9473 | Antigen | Non-allergen | Non-toxin | 11 |
| ATYYFAELEVAVKH | PD118/2017 (India) | 0.7559 | Antigen | Allergen | Non-toxin | 8.8 | ||
| BoLA-T7 (2–15) | 2–15 | TSTGESADPVTTTV | Strain O/ME-SA/Ind, PD287/2015, PD118/2017, PD73/2017, IRN/72/2009, BHU/3/2009, O/OMN/7/2001, O/KUW/3/97, INDIA 2013, Depok 2, Sukabumi 1, 2 | 0.8275 | Antigen | Allergen | Non-toxin | 12 |
| TSAGESADPVTTTV | MYA/13/2017 (Myanmar) | 0.7412 | Antigen | Non-allergen | Non-toxin | 11 | ||
| TSAGESADPVTATV | ISA/GRESIK/2022, ISA/BANJARNEGARA/2022, INDONESIA 2022 | 0.7250 | Antigen | Non-allergen | Non-toxin | 13 | ||
| TSTGESADPVTATV | Bandung 1, 3, 4, 5, 8, 9, Palembang 8 | 0.8113 | Antigen | Allergen | Non-toxin | 14 |
BoLA = Bovine leukocyte antigen.
B-cell epitope prediction indicated that Sukabumi 1, Sukabumi 2, and Depok 2 differed from ISA/2022 and ISA/Banjarnegara/Gresik by an alanine-to-threonine substitution at position 13 within the 6–30 epitope region. These profiles were otherwise consistent with other FMDV serotype O database isolates. Notably, Palembang 8 exhibited a glutamic acid–to–lysine substitution at position 196 within the 192–205 epitope region, distinguishing it from Ind/2022, ISA/Banjarnegara/Gresik, and other reference isolates. Peptides spanning positions 93–107 were predicted to be non-antigenic across all samples, whereas other regions demonstrated antigenic potential. Several peptides showed predicted allergenicity, while all remained non-toxic (Table 7).
Table 7. Prediction of VP1 binding to B cells among collected samples and database.
| Position | Peptide sequence | Databases and code samples | Antigenicity prediction score | Antigenicity | Allergenicity | Toxicity |
|---|---|---|---|---|---|---|
| 6–30 | ESADPVTTTVENYGGETQVQRRQHT | Strain O/ME-SA/Ind, MYA/13/2017 (Myanmar), PD287/2015 (India), PD118/2017, PD73/2017 (India), IRN/72/2009 (Iran), BHU/3/2009 (Bhutan), O/OMN/7/2001 (Oman), O/KUW/3/97 (Kuwait), INDIA 2013, Depok 2, Sukabumi 1, 2 | 0.5586 | Antigen | Allergen | Non-toxin |
| ESADPVTATVENYGGETQVQRRQHT | ISA/GRESIK (Indonesia), ISA/BANJARNEGARA (Indonesia), INDONESIA 2022, Bandung 1, 3, 4, 5, 8, 9, Depok 1, Bogor 1, Palembang 1, 2, 3, 4, 6, 7, 8 | 0.5596 | Antigen | Allergen | Non-toxin | |
| 41–58 | KVTPKDQINVLDLMQTPA | Strain O/ME-SA/Ind, MYA/13/2017 (Myanmar), PD287/2015 (India), PD118/2017, PD73/2017 (India), IRN/72/2009 (Iran), BHU/3/2009 (Bhutan), O/KUW/3/97 (Kuwait), INDIA 2013, ISA/GRESIK (Indonesia), ISA/BANJARNEGARA (Indonesia), INDONESIA 2022, Depok 1, 2, Sukabumi 1, 2, Bandung 1, 3, 4, 5, 8, 9, Bogor 1, Palembang 1, 2, 3, 4, 6, 7, 8 | 0.8708 | Antigen | Non-allergen | Non-toxin |
| KVTPKDQINVLDLMKTPA | O/OMN/7/2001 (Oman) | 0.7528 | Antigen | Non-allergen | Non-toxin | |
| 93–107 | APETALDNTTNPTAY | Strain O/ME-SA/Ind, O/KUW/3/97 (Kuwait) | 0.2242 | Non-antigen | Allergen | Non-toxin |
| APEAALDNTTNPTAY | MYA/13/2017 (Myanmar), PD287/2015 (India), IRN/72/2009 (Iran), BHU/3/2009 (Bhutan), INDIA 2013 | 0.2399 | Non-antigen | Non-allergen | Non-toxin | |
| APEVALDNTTNPTAY | PD118/2017 (India), PD73/2017 (India) | 0.3417 | Non-antigen | Allergen | Non-toxin | |
| APEKALDNTTNPTAY | O/OMN/7/2001 (Oman) | 0.1378 | Non-antigen | Allergen | Non-toxin | |
| APETALENTTNPTAY | ISA/GRESIK (Indonesia), ISA/BANJARNEGARA (Indonesia), INDONESIA 2022, Depok 1, 2, Sukabumi 1, 2, Bandung 1, 3, 4, 5, 8, 9, Bogor 1, Palembang 1, 2, 3, 4, 6, 7, 8 | 0.3708 | Non-antigen | Allergen | Non-toxin | |
| 133–150 | NCKYGEGAVTNVRGDLQV | Strain O/ME-SA/Ind, MYA/13/2017 (Myanmar), PD287/2015 (India), O/OMN/7/2001 (Oman) | 0.5798 | Antigen | Allergen | Non-toxin |
| NCRYGAGAVTNVRGDLQV | PD118/2017 (India), PD73/2017 (India) | 0.7176 | Antigen | Allergen | Non- toxin | |
| NCKYGESDVTNVRGDLQV | IRN/72/2009 (Iran), BHU/3/2009 (Bhutan) | 0.7300 | Antigen | Allergen | Non-toxin | |
| DCKYGESAVTNVRGDLQV | O/KUW/3/97 (Kuwait) | 0.7244 | Antigen | Allergen | Non-toxin | |
| NCKYGENNVPNVRGDLQV | INDIA 2013 | 0.6021 | Antigen | Allergen | Non-toxin | |
| NCKYGEGAVANVRGDLQV | ISA/GRESIK (Indonesia), ISA/BANJARNEGARA (Indonesia), INDONESIA 2022, Depok 1, 2, Sukabumi 1, 2, Bandung 1, 3, 4, 5, 8, 9, Bogor 1, Palembang 1, 2, 3, 4, 6, 7, 8 | 0.5417 | Antigen | Allergen | Non-toxin | |
| 192–205 | AIHPEQARHKQKIV | Strain O/ME-SA/Ind, MYA/13/2017 (Myanmar), PD287/2015 (India), PD118/2017, PD73/2017 (India), ISA/GRESIK (Indonesia), ISA/BANJARNEGARA (Indonesia), INDONESIA 2022, Depok 1, 2, Sukabumi 1, 2, Bandung 1, 3, 4, 5, 8, 9, Bogor 1, Palembang 1, 2, 3, 4, 6, 7 | 0.6571 | Antigen | Non-allergen | Non-toxin |
| AIHPSQARHKQKIV | IRN/72/2009 (Iran), BHU/3/2009 (Bhutan) | 0.7821 | Antigen | Non-allergen | Non-toxin | |
| AIHPSEARHKQKIV | O/OMN/7/2001 (Oman), O/KUW/3/97 (Kuwait), INDIA 2013 | 0.8339 | Antigen | Non-allergen | Non-toxin | |
| AIHPKQARHKQKIV | Palembang 8 | 0.7958 | Antigen | Allergen | Non-toxin |
Prediction of VP1 protein–ligand binding sites
Protein–ligand binding site analysis of
Table 8. Analysis of the ligand-binding site of VP1 protein among collected samples and database.
| No. | Sample | Pocket No. | Residues | Score | Rank |
|---|---|---|---|---|---|
| 1 | O/ME_SA/Ind | 1 | 85_ASN, 87_THR, 98_LEU, 99_ASP, 101_THR, 105_THR, 168_LYS, 169_ALA, 170_THR | 1.92 | 1 |
| 2 | 50_VAL, 89_VAL, 90_PRO, 91_ASN, 92_GLY, 93_ALA, 163_ASN, 165_GLY, 166_ALA | 1.79 | 2 | ||
| 3 | 86_LEU, 88_TRP, 167_ILE | 0.97 | 3 | ||
| 2 | MYA/13/2017 | 1 | 85_ASN, 87_THR, 98_LEU, 99_ASP, 101_THR, 105_THR, 168_LYS, 169_ALA, 170_THR | 1.92 | 1 |
| 2 | 50_VAL, 89_VAL, 90_PRO, 91_ASN, 92_GLY, 93_ALA, 163_ASN, 165_GLY, 166_ALA | 1.76 | 2 | ||
| 3 | 78_VAL, 80_VAL, 86_LEU, 88_TRP, 115_LEU, 167_ILE | 0.97 | 3 | ||
| 3 | PD287/2015 | 1 | 71_TYR, 123_HIS, 125_VAL, 127_ALA, 130_TYR, 162_PHE | 2.41 | 1 |
| 2 | 50_VAL, 89_VAL, 90_PRO, 92_GLY, 93_ALA, 95_GLU, 98_LEU, 163_ASN, 165_GLY, 166_ALA | 2.21 | 2 | ||
| 3 | 86_ASN, 87_THR, 99_ASP, 101_THR, 168_LYS, 169_ALA, 170_THR | 1.31 | 3 | ||
| 4 | 52_ASP, 53_LEU, 72_TYR, 164_TYR | 0.81 | 4 | ||
| 4 | PD118/2017 | 1 | 71_TYR, 123_HIS, 125_VAL, 127_ALA, 130_TYR, 134_CYS, 160_PRO, 161_SER, 162_PHE | 3.13 | 1 |
| 2 | 132_GLY, 133_ASN, 135_ARG, 138_ALA, 141_VAL, 142_THR, 153_GLN, 156_ALA, 157_ARG, 158_THR | 2.30 | 2 | ||
| 3 | 50_VAL, 89_VAL, 90_PRO, 93_ALA, 95_GLU, 98_LEU, 163_LYS, 166_ALA | 2.06 | 3 | ||
| 4 | 52_ASP, 53_LEU, 72_TYR, 164_TYR | 0.90 | 4 | ||
| 5 | PD73/2017 | 1 | 71_TYR, 125_VAL, 127_ALA, 130_TYR, 134_CYS | 3.13 | 1 |
| 2 | 132_GLY, 133_ASN, 135_ARG, 138_ALA, 141_VAL, 142_THR, 153_GLN, 156_ALA, 157_ARG, 158_THR | 2.30 | 2 | ||
| 3 | 89_VAL, 90_PRO, 93_ALA, 95_GLU, 98_LEU, 163_LYS, 166_ALA | 2.05 | 3 | ||
| 4 | 52_ASP, 53_LEU, 72_TYR, 164_TYR | 0.90 | 4 | ||
| 6 | IRN/72/2009; BHU/3/2009 | 1 | 85_ASN, 87_THR, 98_LEU, 99_ASP, 101_THR, 105_THR, 168_LYS, 169_ALA, 170_THR | 1.92 | 1 |
| 2 | 50_VAL, 89_VAL, 90_PRO, 91_ASN, 92_GLY, 93_ALA, 163_ASN, 165_GLY, 166_ALA | 1.72 | 2 | ||
| 3 | 78_VAL, 80_VAL, 86_LEU, 88_TRP, 115_LEU, 167_ILE | 0.98 | 3 | ||
| 7 | O/OMN/7/2001 | 1 | 50_VAL, 89_VAL, 90_PRO, 91_ASN, 92_GLY, 93_ALA, 95_GLU, 163_ASN, 165_GLY, 166_ALA | 2.35 | 1 |
| 2 | 85_ASN, 87_THR, 98_LEU, 99_ASP, 101_THR, 105_THR, 168_LYS, 169_ALA, 170_THR | 1.95 | 2 | ||
| 3 | 78_VAL, 80_VAL, 86_LEU, 88_TRP, 115_LEU, 167_ILE | 0.98 | 3 | ||
| 8 | O/KUW/3/97 | 1 | 50_VAL, 89_VAL, 90_PRO, 91_ASN, 92_GLY, 93_ALA, 95_GLU, 163_ASN, 165_GKY, 166_ALA | 2.26 | 1 |
| 2 | 85_ASN, 87_THR, 98_LEU, 99_ASP, 101_THR, 105_THR, 168_LYS, 169_ALA, 170_THR | 2.00 | 2 | ||
| 3 | 78_VAL, 80_VAL, 86_LEU, 88_TRP, 115_LEU, 167_ILE | 1.06 | 3 | ||
| 9 | INDIA 2013 | 1 | 85_ASN, 87_THR, 98_LEU, 99_ASP, 100_ASN, 101_THR, 105_THR, 168_LYS, 169_ALA, 170_THR | 1.71 | 1 |
| 2 | 135_LYS, 139_ASN, 140_ASN, 156_ALA | 1.36 | 2 | ||
| 10 | INDONESIA 2022; ISA/GRESIK; ISA/BANJARNEGARA | 1 | 85_ASN, 87_THR, 98_LEU, 99_GLU, 168_LYS, 169_ALA, 170_THR | 2.11 | 1 |
| 2 | 50_VAL, 89_VAL, 90_PRO, 91_ASN, 92_GLY, 93_ALA, 163_ASN, 165_GLY, 166_ALA | 1.78 | 2 | ||
| 3 | 78_VAL, 80_VAL, 86_LEU, 88_TRP, 115_LEU, 167_ILE | 0.97 | 3 | ||
| 11 | BANDUNG; DEPOK; BOGOR; PALEMBANG | 1 | 71_TYR, 123_HIS, 125_VAL, 127_ALA, 130_TYR, 162_PHE | 2.44 | 1 |
| 2 | 50_VAL, 89_VAL, 90_PRO, 92_GLY, 93_ALA, 95_GLU, 98_LEU, 163_ASN, 165_GLY, 166_ALA | 2.21 | 2 | ||
| 3 | 52_ASP, 72_TYR, 164_TYR | 0.82 | 3 | ||
| 12 | DEPOK; SUKABUMI | 1 | 71_TYR, 123_HIS, 125_VAL, 127_ALA, 130_TYR, 162_PHE | 2.36 | 1 |
| 2 | 50_VAL, 89_VAL, 90_PRO, 92_GLY, 93_ALA, 95_GLU, 98_LEU, 163_ASN, 165_GLY, 166_ALA | 2.16 | 2 | ||
| 3 | 52_ASP, 53_LEU, 72_TYR, 164_TYR | 0.81 | 3 | ||
| 13 | PALEMBANG 8 | 1 | 71_TYR, 123_HIS, 125_VAL, 127_ALA, 130_TYR, 162_PHE | 2.38 | 1 |
| 2 | 50_VAL, 89_VAL, 90_PRO, 92_GLY, 93_ALA, 95_GLU, 98_LEU, 163_ASN, 165_GLY, 166_ALA | 2.18 | 2 | ||
| 3 | 132_GLY, 133_ASN, 138_GLU, 153_GLN, 156_THR, 157_ARG, 158_ALA | 2.10 | 3 | ||
| 4 | 85_ASN, 87_THR, 99_GLU, 101_THR, 168_LYS, 169_ALA, 170_THR | 1.51 | 4 | ||
| 5 | 52_ASP, 53_LEU, 72_TYR, 164_TYR | 0.79 | 5 | ||
| 14 | BANDUNG 8 | 1 | 71_TYR, 122_PRO, 123_HIS, 125_VAL, 127_ALA, 130_TYR, 133_ASN, 134_CYS, 160_PRO, 161_THR, 162_SER, 163_PHE | 3.19 | 1 |
| 2 | 50_VAL, 89_VAL, 90_PRO, 92_GLY, 93_ALA, 95_GLU, 98_LEU, 164_ASN, 165_GLY, 167_ALA | 2.27 | 2 | ||
| 3 | 85_ASN, 87_THR, 99_GLU, 101_THR, 169_LYS, 170_ALA, 171_THR | 1.58 | 3 |
Figure 4. Ligand-binding site prediction and visualization of protein 3D structure using the PrankWeb server. Different colors indicate different pockets. Sample codes: strain O/ME_SA/Ind (1), MYA/13/2017 (2), PD287/2015 (3), PD118/2017 (4), PD73/2017 (5), IRN/72/2009, BHU/3/2009 (6), O/OMN/7/200 (7), O/KUW/3/97 (8), INDIA 2013 (9), ISA/GRESIK, ISA/BANJARNEGARA, INDONESIA 2022 (10), BANDUNG 1, 3, 4, 5, 9, DEPOK 1, BOGOR 1, PALEMBANG 1, 2, 3, 4, 6, 7, 9 (11), DEPOK 2, SUKABUMI 1, 2 (12), PALEMBANG 8 (13), BANDUNG 8 (14).
Molecular docking analysis between VP1 and TLR7
Molecular docking analysis demonstrated variable interaction strength between
Table 9. Prediction of interactions between VP1 protein and TLR7 using amino acid sequences from collected samples and database.
| Sample | Docking score | Confidence score | RMSD | van der Waals energy | Electrostatic energy | Z-score | Cluster size |
|---|---|---|---|---|---|---|---|
| O/ME-SA/Ind | −286.07 | 0.9383 | 19.4 ± 0.1 | −44.1 ± 4.6 | −187.5 ± 19.1 | −1.9 | 14 |
| MYA/13/2017 | −284.87 | 0.9369 | 13.3 ± 0.3 | −49.3 ± 5.0 | −208.8 ± 39.8 | −2.4 | 4 |
| PD287/2015 | −320.49 | 0.9680 | 24.8 ± 0.4 | −63.0 ± 8.9 | −169.9 ± 25.6 | −1.3 | 68 |
| PD118/2017 | −319.86 | 0.9676 | 0.8 ± 0.5 | −70.9 ± 0.5 | −128.5 ± 29.8 | −1.7 | 5 |
| PD73/2017 | −311.30 | 0.9618 | 0.7 ± 0.4 | −74.9 ± 3.5 | −138.9 ± 23.4 | −2.0 | 8 |
| IRN/72/2009; BHU/3/2009 | −292.93 | 0.9458 | 10.0 ± 0.5 | −52.1 ± 11.4 | −175.9 ± 23.5 | −2.2 | 5 |
| O/OMN/7/2001 | −287.50 | 0.9399 | 17.0 ± 0.1 | −42.9 ± 12.5 | −206.1 ± 15.3 | −1.5 | 4 |
| O/KUW/3/97 | −277.89 | 0.9281 | 14.8 ± 0.1 | −49.8 ± 10.6 | −174.2 ± 35.2 | −2.3 | 4 |
| INDIA 2013 | −288.23 | 0.9407 | 10.7 ± 0.2 | −49.8 ± 9.8 | −138.1 ± 28.5 | −1.1 | 6 |
| INDONESIA 2022; ISA/GRESIK; ISA/BANJARNEGARA | −283.24 | 0.9349 | 12.0 ± 0.3 | −45.9 ± 9.0 | −315.8 ± 37.4 | −2.4 | 7 |
| BANDUNG 1, 3, 4, 5, 9; DEPOK 1; BOGOR 1; PALEMBANG 1, 2, 3, 4, 6, 7, 9 | −287.37 | 0.9398 | — | −74.3 ± 3.5 | −135.7 ± 8.8 | −1.7 | 11 |
| DEPOK 2; SUKABUMI 1, 2 | −319.04 | 0.9671 | 0.5 ± 0.3 | −68.3 ± 8.4 | −150.0 ± 27.1 | −1.2 | 4 |
| PALEMBANG 8 | −324.27 | 0.9703 | 24.6 ± 0.1 | −65.4 ± 6.7 | −183.7 ± 18.3 | −1.2 | 65 |
| BANDUNG 8 | −321.75 | 0.9688 | 24.8 ± 0.3 | −64.9 ± 4.5 | −168.6 ± 20.1 | −1.4 | 76 |
RMSD = Root mean square deviation.
Table 10. Residue interactions between VP1 protein and TLR7 among collected samples and database.
| Interaction parameter | A | B | C | D | E | F | G | H | I | J | K | L | M |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of patients | 50 | 50 | 38 | 39 | 31 | 53 | 53 | 55 | 45 | 40 | 39 | 44 | 38 |
| Hydrogen bond (%) | 24.0 | 22.0 | 23.7 | 17.9 | 9.7 | 22.6 | 18.9 | 18.2 | 20.5 | 20.5 | 17.9 | 25.0 | 21.1 |
| Van der Waals (%) | 66.0 | 68.0 | 55.3 | 61.5 | 64.5 | 64.2 | 71.7 | 72.7 | 70.5 | 71.8 | 64.1 | 59.1 | 60.5 |
| π–π stack (%) | 2.0 | 2.0 | 10.5 | 12.8 | 16.1 | 1.9 | 1.9 | 1.8 | 2.3 | 2.6 | 15.4 | 11.4 | 10.5 |
| Ionic (%) | 4.0 | 4.0 | 5.3 | 5.1 | 3.2 | 3.8 | 3.8 | 3.6 | 4.4 | 5.1 | 2.6 | 2.3 | 5.3 |
| π-cation (%) | 2.0 | 2.0 | 2.6 | 2.6 | 3.2 | 3.8 | 1.9 | 1.8 | 2.3 | 0.0 | 0.0 | 2.3 | 2.6 |
A = O/ME-SA/Ind, B = MYA/13/2017 (Myanmar), C = PD287/2015 (India), D = PD118/2017 (India), E = PD73/2017 (India), F = IRN/72/2009 (Iran), BHU/3/2009 (Bhutan), G = O/KUW/3/97 (Kuwait), H = INDIA 2013, I = ISA/1/2022, ISA/Gresik/2022, ISA/Banjarnegara/2022, J = Bandung 1, 3, 4, 5, 9, Depok 1, Bogor 1, Palembang 1, 2, 3, 4, 6, 7, 9, K = Depok 2, Sukabumi 1, Sukabumi 2, L = Palembang 8, M = Bandung 8, TLR7 = Toll-like receptor 7, VP1 = Viral protein 1, aa = Amino acid.
Figure 5. Visualization of docking between the VP1 protein and TLR7 using the HDOCK server, VP1 (green), TLR7 (orange).
DISCUSSION
Persistence of FMD despite vaccination in Indonesia
FMD remains the most damaging viral disease affecting cattle production in Indonesia. Although a national FMD control program has been implemented using an inactivated vaccine containing serotype O strains O/Mya98/XJ/2010, O1 Campos, O1 Manisa, and IND/O/R2/75 (Decree of the Minister of Agriculture of the Republic of Indonesia No. 738/KPTS/PK.300/M/10/2022), FMD continues to circulate within the country. The vaccination campaign has reduced the frequency of clinical disease; however, unregulated animal movement facilitates the ongoing circulation of FMDV strains. In addition, Indonesia’s geographic position increases the risk of incursion of circulating strains from other Asian countries. Consequently, serotype O remains endemic and continues to circulate in Indonesia [6].
Molecular diversity of capsid genes during the 2022–2023 outbreaks
This study presents the first molecular analysis of
Evolutionary mechanisms shaping FMDV diversity
The observed genetic heterogeneity of serotype O FMDV across different regions is likely driven by unrestricted livestock movement. Endemic co-circulation of multiple genotypes may facilitate recombination when closely related strains co-infect a host [17]. As an RNA virus lacking proofreading mechanisms, FMDV evolves rapidly, with mutation rates ranging from 10-3 to 10-5 substitutions per nucleotide site per year [18]. Each viral population exists as a quasispecies comprising genetically and antigenically diverse variants [19]. Because the principal immunogenic region of FMDV is located within the 1D region,
Implications for vaccine effectiveness and disease control
FMDV evolution predominantly occurs through point mutations, and the extent of genetic change influences viral adaptability, transmissibility, and pathogenicity [21, 22]. The emergence of new strains may compromise disease control in endemic regions when vaccine strains fail to provide adequate cross-protection [23]. Moreover, current vaccines do not prevent the establishment of a carrier state, as infected cattle and buffaloes may excrete the virus for extended periods regardless of vaccination status. Although mortality is uncommon, FMD causes severe disease, and vaccination primarily reduces clinical severity rather than preventing infection or halting transmission.
Immunoinformatic insights into VP1 epitope variation
Epitope prediction analysis of
Structural and functional relevance of VP1 binding sites
Previous epitope studies of serotype O have primarily focused on neutralizing domains of
Molecular docking of VP1 with TLR7 and study limitations
Protein–protein docking analysis demonstrated that Palembang 8 showed the lowest docking and confidence scores for interaction between
CONCLUSION
This study provides comprehensive molecular, immunoinformatic, and structural evidence that FMDV serotype O circulating in Indonesia during the 2022–2023 outbreaks is undergoing active genetic and antigenic diversification. Sequence analysis demonstrated reduced nucleotide identity in
These findings have direct implications for FMD control in Indonesia. The observed antigenic drift in
The major strength of this work lies in its integrated approach, combining molecular sequencing, phylogenetic analysis, epitope prediction, antigenicity/allergenicity/toxicity screening, structural pocket mapping, and protein–protein docking. This multi-layered framework enabled functional interpretation of genetic variation beyond descriptive phylogeny and represents the first report linking
This study is limited by its reliance on in silico analyses without experimental validation. Neutralization assays, empirical antigenicity testing, and functional immune assays were not performed. In addition, the number of isolates analyzed, although representative, may not capture the full national diversity of circulating FMDV serotype O.
Future studies should include wet-lab validation of predicted epitopes, vaccine cross-neutralization assays, and longitudinal surveillance to monitor epitope conservancy. Structural refinement using higher-resolution models and experimental confirmation of
Overall, this study demonstrates that FMDV serotype O in Indonesia is evolving through measurable genetic, antigenic, and structural changes with potential consequences for vaccine effectiveness and immune recognition. The findings underscore the need for continuous, integrative surveillance and evidence-based vaccine adaptation to sustain effective FMD control in endemic settings.
DATA AVAILABILITY
The data supporting this study are included in the manuscript. All full sequence VP1 data were deposited in GeneBank with accession numbers PX406137-PX406155.
AUTHORS’ CONTRIBUTIONS
RIA1, ATW, RIA2, NLPIM, and TPP: Drafted the manuscript. RIA1, SS, TPP, and HR: Collected the sample and performed data analysis. RIA1: Extracted RNA, primer design, and gene amplification. RIA1 and TP: Performed epitope and docking analysis. ATW, RIA2, NLPIM, and TPP: Methodology and software validation. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
This research was supported by RIIM Kompetisi Funding from The Indonesia Endowment Fund for Education Agency (LPDP) No: B-1744/II.7.5/FR/11/2022, B-14145/III.11/KS.00.00/11/2022 and Beasiswa Pendidikan Indonesia (BPI) LPDP
REFERENCES
- Li P, Huang S, Zha J, Sun P, Li D, Bao H. Evaluation of immunogenicity and cross-reactive responses of vaccines prepared from two chimeric serotype O foot-and-mouth disease viruses in pigs and cattle. Vet. Res 2022;53(56):1-12. [Google Scholar] | [Crossref]
- Mason P. W, Grubman M. J, Baxt B. Molecular basis of pathogenesis of FMDV. Virus. Res 2003;91:9-32. [Google Scholar] | [Crossref]
- Liu W, Yang B, Wang M, Liang W, Wang H, Yang D. Identification of a conserved conformational epitope in the VP2 protein of foot-and-mouth disease virus. Arch. Virol 2017;162(7):1877-1885. [Google Scholar] | [Crossref]
- Zhu Z, Yang F, Cao W, Liu H, Zhang K, Tian H. The pseudoknot region of the 5′untranslated region is a determinant of viral tropism and virulence of foot-and-mouth disease virus. J. Virol 2019;93(8):e02039-18. [Google Scholar] | [Crossref]
- Abeyratne S. A. E, Amarasekera S. S. C, Ranaweera L. T, Salpadoru T. B, Thilakarathne S. M. N. K, Knowles N. J. Phylogenetic analysis of the VP1 genomic region in foot-and-mouth disease virus serotype O isolates in Sri Lanka reveals the existence of “Srl-97”, a newly named endemic lineage. PLoS One 2018;13(4):e0196491. [Google Scholar] | [Crossref]
- Zainuddin N, Susila E. B, Wibawa H, Daulay R. S. D, Wijayanti P. E, Fitriani D. Genome sequence of a foot-and-mouth disease virus detected in Indonesia in 2022. Microbiol. Resour. Announc 2023;12(2):e0108122. [Google Scholar] | [Crossref]
- Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Paris: WOAH; 2025. [Google Scholar]
- Zhang X, Ma W, Liu B, Shen C, Yang F, Yang Y. Phylogenetic analyses and antigenic characterization of foot-and-mouth disease virus PanAsia lineage circulating in China between 1999 and 2023. Virol. Sin 2024;39(5):747-754. [Google Scholar] | [Crossref]
- Reynisson B, Alvarez B, Paul S, Peters B, Nielsen M. NetMHCpan-4.1 and NetMHCIIpan-4.0:Improved predictions of MHC antigen presentation by concurrent motif deconvolution and integration of MS MHC eluted ligand data. Nucleic. Acids. Res 2020;48(1):449-454. [Google Scholar] | [Crossref]
- Jespersen M. C, Peters B, Nielsen M, Marcatili P. BepiPred-2.0:Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic. Acids. Res 2017;45(1):24-29. [Google Scholar] | [Crossref]
- Doytchinova I. A, Flower D. R. VaxiJen:A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform 2007;8((4)). [Google Scholar] | [Crossref]
- Dimitrov I, Bangov I, Flower D. R. Doytchinova I. AllerTOP v.2 –A server for
in silico prediction of allergens. J. Mol. Model 2014;20(6):2278. [Google Scholar] | [Crossref] - Dimitrov I, Naneva L, Doytchinova I, Bangov I. AllergenFP:Allergenicity prediction by descriptor fingerprints. Bioinformatics 2014;30(6):846-851. [Google Scholar] | [Crossref]
- Gupta S, Kapoor P, Chaudhary K, Gautam A, Kumar R, Raghava G. P. In silico approach for predicting toxicity of peptides and proteins. PLoS One 2013;8(9):e73957. [Google Scholar] | [Crossref]
- Jendele L, Krivak R, Skoda P, Novotny M, Hoksza D. PrankWeb:A web server for ligand-binding site prediction and visualization. Nucleic. Acids. Res 2019;47(2):345-349. [Google Scholar] | [Crossref]
- Li H, Huang E, Zhang Y, Huang S. Y, Xiao Y. HDOCK update for modeling protein–RNA/DNA complex structures. Protein. Sci 2022;31(11):e4441. [Google Scholar] | [Crossref]
- Subramaniam S, Mohapatra J. K, Sahoo N. R, Sahoo A. P, Dahiya S. S, Rout M. Foot-and-mouth disease status in India during the second decade of the twenty-first century (2011–2020). Vet. Res. Commun 2022;46:1011-1022. [Google Scholar] | [Crossref]
- Domingo E, García-Crespo C, Lobo-Vega R, Perales C. Mutation rates, mutation frequencies, and proofreading-repair activities in RNA virus genetics. Viruses 2021;13(9):1882. [Google Scholar] | [Crossref]
- Fish I, Stenfeldt C, Palinski R. M, Pauszek S. J, Arzt J. Into the deep (sequence) of the foot-and-mouth disease virus gene pool:Bottlenecks and adaptation during infection in naïve and vaccinated cattle. Pathogens 2020;9(3):208. [Google Scholar] | [Crossref]
- Hassan A. M, Zaher M. R, Hassanien R. T, Abd-El-Moniem M. I, Habashi A. R, Ibraheem E. M. Molecular detection, phylogenetic analysis and genetic diversity of recently isolated foot-and-mouth disease virus serotype A African topotype, Genotype IV. Virol. J 2022;19(1):1. [Google Scholar] | [Crossref]
- Arzt J, Fish I, Pauszek S. J, Johnson S. L, Chain P. S, Rai D. K. The evolution of a super-swarm of foot-and-mouth disease virus in cattle. PLoS One 2019;14(4):e0210847. [Google Scholar] | [Crossref]
- Dahiya S. S, Subramaniam S, Mohapatra J. K, Rout M, Biswal J. K, Giri P. Foot-and-mouth disease virus serotype O exhibits phenomenal genetic lineage diversity in India during 2018–2022. Viruses 2023;15(7):1529. [Google Scholar] | [Crossref]
- Paton D. J, Sumption K. J, Charleston B. Options for control of foot-and-mouth disease:Knowledge, capability and policy. Philos. Trans. R Soc Lond. B Biol. Sci 2009;364(1530):2657-2667. [Google Scholar] | [Crossref]
- Woldemariyam F, Paeshuyse J. Viral protein 1 (VP1) sequence-based genetic diversity of SAT 2 FMDV circulating in Ethiopia from 1990 to 2015. Vet. Med 2023;14:91-101. [Google Scholar] | [Crossref]
- Elrashedy A, Nayel M, Salama A, Zaghawa A, El-Shabasy R. M, Hasan M. E. Foot-and-mouth disease:Genomic and proteomic structure, antigenic sites, serotype relationships, immune evasion, recent vaccine development strategies, and future perspectives. Vet. Res 2025;56(78):1-29. [Google Scholar] | [Crossref]
- Hussein H. A. M, Walker L. R, Abdel-Raouf U. M, Desouky S. A, Montasser A. K. M, Akula S. M. Beyond RGD:Virus interactions with integrins. Arch. Virol 2015;160(11):2669-2681. [Google Scholar] | [Crossref]
- Medina G. N, Diaz San Segundo F. Virulence and immune evasion strategies of FMDV:Implications for vaccine design. Vaccines 2024;12(9):1071. [Google Scholar] | [Crossref]
- Babiuk S, Horseman B, Zhang C, Bickis M, Kusalik A, Schook L. B. BoLA class I allele diversity and polymorphism in a herd of cattle. Immunogenetics 2007;59(2):167-176. [Google Scholar] | [Crossref]
- Zhao Q, Li F, Huang S, Xing X, Sun Y, Li P. Remapping the spatial distribution of neutralizing sites and their immunodominance on the capsid of different topotypes of FMDV serotype O by site-directed competitive ELISA for detection of neutralizing antibodies. Microbiol. Spectr 2025;13(16):e03344-24. [Google Scholar] | [Crossref]
- He Y, Li K, Cao Y, Sun Z, Li P, Bao H. Structures of foot-and-mouth disease virus with neutralizing antibodies derived from recovered natural host reveal a mechanism for cross-serotype neutralization. PLoS. Pathog 2021;17(4):1009507. [Google Scholar] | [Crossref]
- Dong H, Liu P, Bai M, Wang K, Feng R, Zhu D. Structural and molecular basis for foot-and-mouth disease virus neutralization by two potent protective antibodies. Protein. Cell 2022;13:446-453. [Google Scholar] | [Crossref]
- Momtaz S, Rahman A, Sultana M, Hossain M. A. Evolutionary analysis and prediction of peptide vaccine candidates for foot-and-mouth disease virus types A and O in Bangladesh. Evol. Bioinform. Online 2014;10:187-196. [Google Scholar] | [Crossref]
- Yun T, Liu G. Q, Leng Q. W, Guo H. C, Liu Z. X, Zhang J. N, Xie Q. G. Prediction of secondary structure and B-cell epitope for capsid protein of FMDV. Chin. J. Vet. Sci. Technol 2004;34:23-29. [Google Scholar] | [Crossref]
- Aggarwal N, Zhang Z, Cox S, Statham R, Alexandersen S, Kitching R. P, Barnett P. V. Experimental studies with foot-and-mouth disease virus, strain O, responsible for the 2001 epidemic in the United Kingdom. Vaccine 2002;20((19–20)):2508-2515. [Google Scholar] | [Crossref]
- Zhang Y, Liu T, Wang J, Zou B, Li L, Yao L. Cellinker:A platform of ligand–receptor interactions for intercellular communication analysis. Bioinformatics 2021;37:1-8. [Google Scholar] | [Crossref]
- Wang H. Prediction of protein–ligand binding affinity via deep learning models. Brief. Bioinform 2024;25(2):1-17. [Google Scholar] | [Crossref]
- Macalino S. J, Gosu V, Hong S, Choi S. Role of computer-aided drug design in modern drug discovery. Arch. Pharm. Res 2015;38(9):1686-1701. [Google Scholar] | [Crossref]
- Anderson A. C. The process of structure-based drug design. Chem Biol 2003;10(9):787-797. [Google Scholar] | [Crossref]
- Dominguez C, Boelens R, Bonvin A. M. HADDOCK:A protein–protein docking approach based on biochemical or biophysical information. J. Am. Chem. Soc 2003;125(7):1731-1737. [Google Scholar] | [Crossref]