


ABSTRACT
Ovarian hypofunction is a major reproductive disorder in dairy cows and contributes substantially to reduced fertility, prolonged days open, and economic losses. Increasing evidence indicates that this condition is not solely an ovarian problem but part of a broader immunometabolic disturbance that also affects uterine health during the postpartum transition period. Negative energy balance after calving leads to elevated circulating metabolites such as non-esterified fatty acids and β-hydroxybutyrate, which trigger oxidative stress and inflammatory signaling. These changes impair hypothalamic–pituitary–ovarian activity, suppress steroidogenesis, and delay follicular development. At the same time, metabolic stress weakens uterine immune defense, slows uterine involution, and increases susceptibility to endometritis, creating a reciprocal cycle in which uterine inflammation further inhibits ovarian reactivation. This narrative review synthesizes current knowledge on the bidirectional interactions between ovarian function and uterine health from an immunometabolic perspective. A structured literature search of major scientific databases was conducted, focusing on peer-reviewed studies addressing postpartum metabolism, immune responses, reproductive physiology, and non-hormonal therapeutic approaches in dairy cows. The review integrates endocrine, metabolic, and inflammatory mechanisms into a unified framework explaining how immune–metabolic imbalance disrupts reproductive homeostasis. Within this framework, uterine flushing therapy is discussed as a practical non-hormonal intervention aimed at restoring uterine conditions rather than directly inducing ovulation. By removing inflammatory exudates, reducing endotoxin load, improving endometrial perfusion, and supporting immune resolution, flushing may indirectly promote normalization of hormonal signaling and ovarian activity. Field evidence suggests that, when combined with appropriate nutritional and herd management strategies, this approach can improve pregnancy outcomes and reduce reliance on repeated hormonal treatments. Overall, adopting an immunometabolic management strategy that integrates metabolic monitoring, uterine health assessment, and targeted supportive interventions offers a promising pathway to enhance reproductive efficiency and sustainability in modern dairy production systems. Further standardized field trials and biomarker-guided protocols are needed to validate these approaches and facilitate their wider on-farm implementation.
Keywords: biomarkers, dairy cows, immunometabolism, negative energy balance, ovarian hypofunction, reproductive efficiency, sustainable agricultural production, transition period, uterine flushing therapy.
INTRODUCTION
Reproductive issues in dairy cattle directly impact production efficiency and farm profitability [1]. Conditions such as anestrus, repeat breeding, and delayed conception are especially prevalent during early lactation, when physiological and metabolic demands are highest [2]. Globally, ovarian hypofunction (OH) affects approximately 10%–30% of postpartum dairy cows, with higher prevalence in intensive production systems and during early lactation. Regional studies in developing dairy sectors report rates exceeding 35% under suboptimal nutritional and management conditions [3]. These disorders prolong the calving interval, reduce conception rates, and increase the costs of insemination and reproductive care, ultimately lowering the economic efficiency of dairy production systems [4].
OH is a major contributor to reproductive disorders in dairy cows, characterized by reduced ovarian activity that prevents dominant follicle development and normal ovulation [5]. This condition is closely associated with hormonal imbalance and metabolic disturbances during the transition period, when energy requirements rise sharply to support the onset of lactation [6]. During this phase, cows frequently experience negative energy balance (NEB), leading to excessive fat mobilization and increased circulating concentrations of non-esterified fatty acids (NEFA) and β-hydroxybutyrate (BHBA) [7, 8].
Beyond its direct effects on ovarian function, metabolic imbalance also has profound consequences for uterine health [9]. Reduced estrogen concentrations associated with OH decrease myometrial contractility and impair uterine defense mechanisms, increasing the risk of retained lochia, delayed uterine involution, and subclinical endometritis [10]. Economically, reproductive inefficiency linked to OH and uterine inflammation can result in losses ranging from tens to several hundred US dollars per affected cow per lactation, mainly due to extended days open, increased veterinary interventions, and reduced lifetime productivity [11, 12].
From a contemporary physiological perspective, immunometabolism provides a unifying framework for maintaining reproductive homeostasis by linking metabolic status with immune regulation [13]. Metabolic stress and postpartum inflammation can disrupt immune signaling in both the uterus and ovaries [14], making correction of this immune–metabolic imbalance essential for restoring reproductive function in dairy cows [15].
One non-hormonal strategy receiving increasing attention is uterine flushing therapy. This procedure involves flushing the uterine lumen with physiological or immunonutritional solutions to remove inflammatory exudates, improve endometrial perfusion, and modulate local immune responses [16]. Unlike conventional hormonal treatments that primarily induce ovulation, uterine flushing targets the uterine environment itself, with improvement in ovarian activity considered a secondary consequence of restored uterine–ovarian communication. This approach is particularly attractive for on-farm application because of its practicality and minimal interference with systemic hormonal balance [17].
Despite extensive research on postpartum reproductive disorders in dairy cows, most studies have examined OH, uterine disease, metabolic stress, and immune responses as separate entities. Existing reviews typically focus on endocrine control of ovarian activity, clinical aspects of endometritis, or nutritional management of transition cows, but rarely integrate these components into a unified physiological framework. In particular, the mechanistic links through which metabolic stress and immune activation simultaneously influence uterine recovery and ovarian reactivation remain insufficiently synthesized. Furthermore, while uterine flushing therapy has been reported as a practical non-hormonal intervention for improving uterine health, its role is often discussed primarily from a mechanical or clinical standpoint, with limited emphasis on its potential immunometabolic effects or its indirect influence on ovarian function. The lack of an integrative perspective combining metabolic biomarkers, immune signaling pathways, uterine–ovarian crosstalk, and supportive non-hormonal interventions represents a significant knowledge gap. Addressing this gap is essential for developing physiologically oriented and sustainable reproductive management strategies that move beyond symptom-based hormonal treatments toward mechanism-driven herd-level approaches.
Therefore, this narrative review aims to provide a comprehensive synthesis of current evidence on the bidirectional interactions between ovarian function and uterine health in dairy cows within an immunometabolic framework. Specifically, the review seeks to (i) summarize the endocrine, metabolic, and inflammatory mechanisms underlying postpartum OH and uterine dysfunction; (ii) elucidate how metabolic stress and immune dysregulation mediate crosstalk between the uterus and ovaries during the transition period; (iii) evaluate the biological rationale, physiological effects, and practical applications of uterine flushing therapy as a complementary non-hormonal intervention; and (iv) highlight the role of immunometabolic biomarkers and supportive management strategies in improving reproductive efficiency. By integrating these aspects, the review intends to present a coherent conceptual model linking metabolic status, immune regulation, and reproductive performance, thereby supporting evidence-based and sustainable approaches to fertility management in modern dairy production systems.
REVIEW METHODOLOGY
Study design
This study was conducted as a narrative review aimed at synthesizing current evidence on the immunometabolic links between OH and uterine health in dairy cows, with particular emphasis on non-hormonal flushing therapy as a complementary intervention. A narrative approach was selected to enable mechanistic integration of endocrine, immune, and metabolic pathways that are often examined separately in experimental and clinical studies.
Literature search strategy
A structured literature search was performed using the PubMed, Scopus, and Web of Science databases. The search included peer-reviewed articles published between 2000 and December 2025. The following keywords and Boolean combinations were used: “ovarian hypofunction,” “postpartum anestrus,” “uterine health,” “endometritis,” “negative energy balance,” “immunometabolism,” “non-esterified fatty acids,” “β-hydroxybutyrate,” and “uterine flushing therapy.”
Study selection criteria
The inclusion criteria were as follows: (i) original research articles, clinical trials, and relevant review papers; (ii) studies conducted in dairy cows; (iii) investigations evaluating ovarian function, uterine health, metabolic status, or immune responses during the postpartum period; and (iv) studies reporting outcomes related to reproductive performance or uterine recovery. Exclusion criteria included studies involving non-dairy species, non-postpartum animals, case reports lacking diagnostic criteria, and articles without relevance to reproductive physiology or immunometabolic mechanisms.
Definitions and diagnostic standards
OH was defined as the absence of ovulation or dominant follicle development beyond 40–50 days postpartum, characterized by small inactive ovaries on ultrasonography (<8 mm follicles), low circulating progesterone concentrations (<1 ng/mL), and absence of a functional corpus luteum. Subclinical endometritis was defined based on cytological thresholds of ≥5–10% polymorphonuclear neutrophils (PMNs) at 21–35 days postpartum or ≥3–5% PMNs after 35 days postpartum in the absence of clinical signs. Uterine dysfunction included delayed involution beyond 45 days postpartum, persistent intrauterine fluid, or ultrasonographic evidence of inflammatory changes.
Outcome measures
The effectiveness of uterine flushing therapy was evaluated using standardized reproductive outcomes, including pregnancy rate, interval to first estrus, days open, and indicators of uterine recovery, such as cytological and ultrasonographic improvement. Comparisons with hormonal or nutritional interventions were based on reported clinical outcomes and mechanistic relevance rather than formal meta-analysis.
Selection and evaluation of immunometabolic biomarkers
Immunometabolic biomarkers were selected based on their documented association with postpartum metabolic stress, inflammation, and reproductive outcomes. These included metabolic indicators (NEFA, BHBA, glucose, and IGF-1), inflammatory markers (haptoglobin and ceruloplasmin), and cytokines (IL-1β, IL-6, IL-8, and TNF-α). Conflicting findings were interpreted in relation to differences in sampling time, parity, production system, and disease severity.
Data synthesis approach
Evidence was synthesized using a thematic and mechanistic framework. Findings were classified into endocrine regulation, metabolic stress, immune activation, and therapeutic intervention domains. This structured synthesis enabled integration of bidirectional ovary–uterus interactions within an immunometabolic context.
OVARIAN PHYSIOLOGY AND HORMONAL REGULATION
In dairy cows, ovarian function is regulated through coordinated interactions among the hypothalamic–pituitary–ovarian (HPO) axis, metabolic signals, and the animal’s physiological status [18]. The ovaries control estrous cyclicity, ovulation, and steroid hormone production, all of which are essential for fertility [19]. This section focuses on regulatory mechanisms relevant to postpartum ovarian recovery and dysfunction rather than providing a detailed description of estrous cycle phases.
Under normal postpartum conditions, ovarian cyclicity typically resumes within 20–40 days after calving, marked by dominant follicle emergence and first ovulation. In well-managed herds, regular luteal activity and estrous cycles are generally re-established by 40–60 days postpartum [20]. Delays beyond this period represent a significant deviation from normal physiology and are strongly associated with OH, NEB, and concurrent uterine inflammation, as discussed in later immunometabolic sections [21].
Follicle-stimulating hormone (FSH) initiates follicular development by stimulating a cohort of antral follicles [22]. Selection of the dominant follicle is accompanied by increased estradiol (E2) and inhibin secretion, which suppresses further FSH release [23]. Rising estradiol concentrations trigger the preovulatory luteinizing hormone (LH) surge, leading to ovulation and corpus luteum (CL) formation [24]. The CL secretes progesterone (P4), which supports luteal function and prepares the uterus for embryo implantation [25]. Physiologically, circulating progesterone concentrations exceed approximately 1 ng/mL during the functional luteal phase, whereas estradiol peaks during follicular dominance.
Postpartum OH is characterized by deviations from this physiological pattern, including persistently low estradiol concentrations, reduced LH pulse frequency, attenuated or absent LH surges, and delayed or insufficient progesterone secretion. These changes reflect impaired follicular growth and luteal insufficiency and frequently coexist with metabolic and inflammatory disturbances typical of early lactation [26].
Metabolic status plays a pivotal role in modulating ovarian responsiveness [27]. During early lactation, dairy cows commonly experience NEB due to increased energy demands for milk production [28]. This condition is associated with reduced circulating insulin and insulin-like growth factor-1 (IGF-1), both of which are critical for granulosa cell proliferation and estradiol synthesis [29]. Altered leptin signaling during NEB further compromises hypothalamic gonadotropin release, impairing follicular maturation [30]. These metabolic signals form a key link between nutrition and reproductive endocrinology and are further elaborated in Section 4 on immunometabolic ovary–uterus interactions.
Parity and genetic background also influence postpartum ovarian recovery. Compared with primiparous cows, multiparous high-producing cows often experience deeper and more prolonged NEB, increasing susceptibility to delayed ovulation. Likewise, high-yielding breeds such as Holstein–Friesian cattle are more vulnerable to metabolic stress–induced ovarian dysfunction than dual-purpose or indigenous breeds, which typically exhibit greater metabolic resilience [31].
Disruption of metabolic–endocrine balance ultimately leads to OH, defined by reduced follicular activity or failure to ovulate [32]. Oxidative stress and low-grade systemic inflammation, common during the postpartum period, play central roles in this process [33]. Excessive reactive oxygen species (ROS) production damages granulosa cells and suppresses FSH and LH receptor expression [34]. In parallel, proinflammatory cytokines such as TNF-α and IL-6 interfere with steroidogenesis and promote follicular atresia, linking immune activation to endocrine suppression [35]. Elevated NEFA and BHBA further exacerbate ovarian dysfunction through direct cytotoxic effects on oocytes and somatic ovarian cells, reinforcing the immunometabolic mechanisms discussed in subsequent sections [36].
PATHOPHYSIOLOGY OF OH
OH is one of the most prevalent reproductive disorders affecting dairy cows during the early postpartum period [37]. Clinically, it manifests as prolonged anestrus and ovulation failure resulting from impaired endocrine signaling, metabolic imbalance, and ovarian follicular inactivity [38]. Rather than representing an isolated ovarian disorder, OH reflects a multisystem disturbance involving neuroendocrine regulation, energy metabolism, immune activation, and oxidative stress [39].
OH is typically diagnosed in cows that fail to resume ovarian cyclicity beyond 40–60 days postpartum in the absence of overt uterine pathology [40]. Practical assessment relies on repeated transrectal ultrasonography showing the absence of a functional CL, small or static follicles (<8 mm), and persistently low circulating progesterone concentrations (<1 ng/mL) [41, 42]. These indicators help differentiate OH from other postpartum reproductive conditions and align pathophysiological mechanisms with field-level diagnostic tools.
Endocrine dysregulation of the HPO axis
OH can be classified into hypogonadotropic and normogonadotropic forms [43]. In the hypogonadotropic type, reduced gonadotropin-releasing hormone (GnRH) secretion from the hypothalamus decreases LH and FSH release from the anterior pituitary [44]. Consequently, follicular growth fails to reach dominance and ovulation does not occur [45].
In contrast, the normogonadotropic type is characterized by apparently normal circulating gonadotropin concentrations but diminished ovarian responsiveness, often due to altered LH/FSH receptor expression or impaired intracellular signaling within follicular cells [46]. This distinction is clinically important because both forms may show similar ultrasonographic findings but differ in responsiveness to hormonal or non-hormonal therapeutic interventions [47].
Metabolic pathways and NEB
A central driver of postpartum OH is NEB associated with early lactation [48]. High energy demands for milk synthesis reduce circulating glucose, insulin, and IGF-1 levels, all of which are essential modulators of ovarian follicular growth. Reduced insulin and IGF-1 signaling suppress hypothalamic GnRH release and decrease LH pulse frequency, preventing follicles from reaching the ovulatory stage [49].
Blood metabolite profiling provides practical insight into these mechanisms [50]. Cows with OH commonly exhibit elevated NEFA and BHBA concentrations along with reduced glucose and cholesterol levels, the latter being a key precursor for steroid hormone synthesis [51, 52]. Excessive NEFA and BHBA exert lipotoxic effects on oocytes and granulosa cells, promoting apoptosis and further impairing ovarian activity [53].
Inflammatory and oxidative stress mechanisms
Oxidative stress and low-grade systemic inflammation during the periparturient period contribute substantially to ovarian dysfunction [54]. Increased ROS production damages granulosa cells, reduces FSH receptor expression, and disrupts steroidogenesis [55]. Proinflammatory cytokines such as TNF-α and interleukin-6 (IL-6) also interfere with follicular development by suppressing aromatase activity, reducing estradiol synthesis, and impairing ovulation [56, 57]. These immunometabolic interactions provide a mechanistic bridge between metabolic imbalance and endocrine suppression, reinforcing the value of integrating metabolic and inflammatory biomarkers into reproductive assessment strategies.
Morphological and management-related factors
Morphologically, ovaries affected by hypofunction are typically smaller, with smooth surfaces and absence of dominant follicles [43]. Developing follicles often undergo atresia characterized by granulosa cell degeneration, reduced vascularization, and follicular collapse [44]. The CL, when present, is usually inactive or regressing, resulting in inadequate progesterone production [45]. The resulting hypoestrogenic and hypoprogesteronic environment limits uterine stimulation and may delay postpartum endometrial regeneration [46].
Management-related stressors, including heat stress, overcrowding, inadequate ventilation, and high stocking density, can exacerbate NEB and elevate cortisol levels. These stressors further suppress reproductive hormone secretion and increase the risk of prolonged postpartum anestrus, particularly in high-producing dairy herds.
Figure 1 summarizes the integrated endocrine, metabolic, and inflammatory mechanisms underlying OH in early lactation dairy cows.
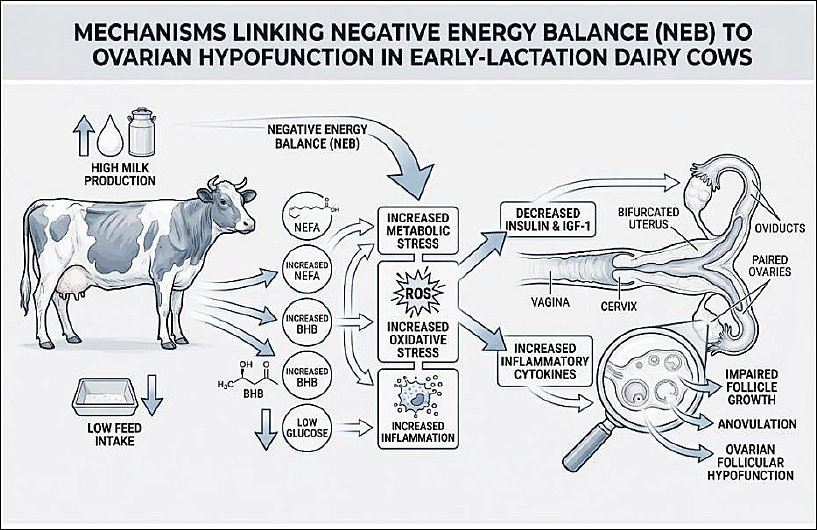
Figure 1. Pathways linking negative energy balance to ovarian hypofunction in early lactation dairy cows. This schematic illustration was generated using artificial intelligence tools (ChatGPT 5.2) and subsequently modified by the authors.
IMMUNOMETABOLIC INTERACTIONS BETWEEN THE OVARY AND UTERUS
Immunometabolism refers to the integrated regulation of metabolic processes, immune function, and endocrine signaling in maintaining physiological homeostasis, including reproductive performance [58]. In dairy cows, immunometabolic balance serves as a central coordinating mechanism linking uterine recovery with ovarian reactivation during the postpartum transition period [59].
Immunometabolic interactions can be conceptualized as a sequential yet bidirectional process in which metabolic stress initially disrupts uterine immune homeostasis, whereas unresolved uterine inflammation subsequently feeds back to impair ovarian function through systemic inflammatory and endocrine signaling [60]. This framework provides a unifying synthesis of mechanisms that have often been described separately in previous reviews. Table 1 illustrates the reciprocal relationships among metabolic, immune, and hormonal systems in dairy cows during the transition period [61–78].
Excessive mobilization of adipose tissue during NEB results in elevated circulating concentrations of NEFA and BHBA [61]. These metabolites function not only as indicators of energy deficit but also as active metabolic triggers that directly modulate immune cell activity [62]. Prolonged exposure to elevated NEFA and BHBA concentrations induces oxidative stress and activates nuclear factor kappa B (NF-κB) signaling in macrophages and endometrial epithelial cells [63]. Consequently, NF-κB activation promotes overproduction of proinflammatory cytokines, including tumor necrosis factor-alpha (TNF-α), interleukin-1β (IL-1β), and IL-6 [64].
Although this inflammatory response is essential for postpartum uterine defense, sustained cytokine release exerts endocrine-disrupting effects that suppress ovarian steroidogenesis, primarily through downregulation of aromatase activity and FSH receptor expression in granulosa cells. This cause–effect cascade ultimately impairs follicular dominance, ovulation, and luteal function [65].
Uterine inflammation, manifesting as clinical or subclinical endometritis, further amplifies ovarian dysfunction through well-defined inflammatory crosstalk mechanisms [66]. Bacterial infection and retained postpartum debris promote the release of lipopolysaccharide (LPS) from Gram-negative bacteria [67]. Systemic translocation of LPS represents a critical mechanistic link, as LPS binds to Toll-like receptor 4 (TLR4) expressed on ovarian granulosa and theca cells, activating NF-κB and p38 mitogen-activated protein kinase pathways [68]. This sequence establishes a direct causal pathway in which uterine inflammation precedes and induces ovarian suppression rather than merely coexisting with it [69]. Furthermore, LPS inhibits expression of steroidogenic acute regulatory protein and cytochrome P450 aromatase (CYP19A1), reducing estradiol synthesis and preventing dominant follicle development [70].
Beyond NEFA and BHBA, additional immunometabolic mediators integrate metabolic status with uterine–ovarian signaling. Adiponectin, an adipokine whose concentration declines during NEB, enhances insulin sensitivity and supports ovarian steroidogenesis; thus, reduced adiponectin levels represent a mechanistic link between metabolic stress and increased inflammatory susceptibility [71]. Cortisol, elevated during metabolic or environmental stress, exerts dual effects by transiently suppressing uterine inflammation while chronically inhibiting gonadotropin secretion and ovarian responsiveness [72]. Acute-phase proteins such as haptoglobin and serum amyloid A serve as systemic biomarkers of immunometabolic imbalance and are consistently associated with delayed resumption of ovarian cyclicity and reduced fertility [73].
At the intracellular level, integration of metabolic and immune signals is mediated through the AMP-activated protein kinase (AMPK)– mechanistic target of rapamycin (mTOR) axis [74]. Activation of mTOR during inflammatory or nutrient-excess states promotes proinflammatory protein synthesis, whereas AMPK activation under energy-deficient conditions conserves cellular energy and restrains inflammatory signaling [75]. Disruption of the AMPK–mTOR balance in uterine and ovarian tissues compromises endometrial repair, granulosa cell survival, and ovulatory capacity, reinforcing a self-perpetuating immunometabolic loop between these organs [76].
The uterine microbiota further modulate these interactions. Under healthy conditions, commensal microbial communities support mucosal integrity and immune tolerance [77]. Postpartum dysbiosis, characterized by overgrowth of pathogenic bacteria such as Escherichia coli and Trueperella pyogenes, sustains chronic inflammation and increases systemic exposure to LPS, thereby intensifying OH through persistent immunometabolic signaling [78].
An integrative schematic summarizing the cause–effect relationships among metabolic stress, immune activation, and endocrine disruption linking uterine inflammation and OH is presented in Figure 2 to enhance conceptual clarity.
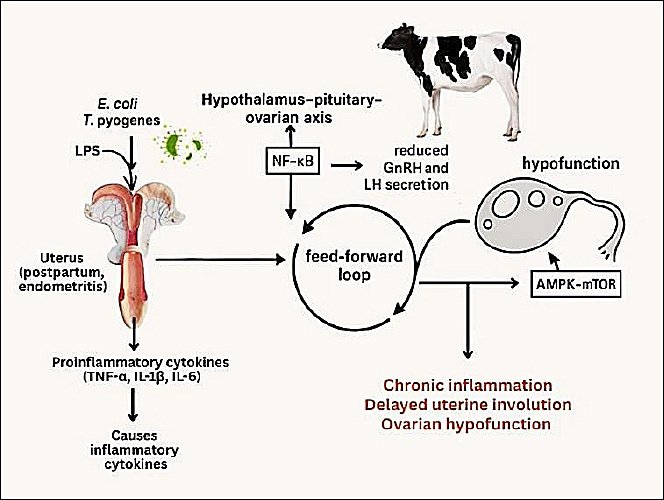
Figure 2. Bidirectional immunometabolic crosstalk between the uterus and ovary in dairy cows with endometritis. This schematic illustration was generated using artificial intelligence tools (ChatGPT 5.2) and subsequently modified by the authors.
Table 1. Immunometabolic interactions between the ovary and uterus during the transition period in female dairy cows.
| Components/Factors | Immunometabolic mechanisms | Impact on the uterus | Impact on the ovaries | Reproductive implications | References |
|---|---|---|---|---|---|
| Negative energy balance | Excessive adipose tissue mobilization elevates circulating NEFA and BHBA levels, which act as active metabolic signals that modulate immune cell function and activate NF-κB pathways | Induces oxidative stress and NF-κB activation in endometrial epithelial cells and macrophages, leading to increased production of IL-1β, IL-6, and TNF-α and delayed uterine immune homeostasis | Proinflammatory cytokines downregulate aromatase (CYP19A1) activity and FSH receptor expression in granulosa cells, suppressing estradiol synthesis | Impaired follicular dominance, delayed ovulation, and reduced luteal function | [61–65] |
|
| |||||
| Postpartum uterine inflammation (endometritis) | Gram-negative bacterial infection releases lipopolysaccharide (LPS), which translocates systemically and activates TLR4-dependent inflammatory signaling | Sustains chronic endometrial inflammation and delays uterine involution and tissue repair | LPS–TLR4 interaction activates NF-κB and p38 mitogen-activated protein kinase pathways in granulosa and theca cells, suppressing steroidogenic acute regulatory and CYP19A1 expression and inhibiting steroidogenesis | Follicular atresia, ovulation failure, and prolonged anestrus | [66–70] |
|
| |||||
| Activation of proinflammatory cytokines (TNF-α, IL-1β, and IL-6) | NF-κB-mediated cytokine overproduction links metabolic stress with immune and endocrine dysregulation | Increases vascular permeability and leukocyte infiltration, promoting chronic low-grade uterine inflammation | Direct suppression of ovarian steroidogenesis and exacerbation of ovarian hypofunction | The reproductive environment becomes unfavorable for conception | [63–65, 69] |
|
| |||||
| AMPK–mTOR signaling axis | Imbalance between AMPK activation during energy deficiency and mTOR activation during inflammation or nutrient-excess amplifies immunometabolic dysregulation | Excessive mTOR activation promotes the synthesis of proinflammatory proteins and impairs endometrial regeneration | Reduced AMPK activity disrupts granulosa cell metabolism, survival, and follicular proliferation | Estrous cycle irregularities and delayed uterine recovery | [74–76] |
|
| |||||
| Uterine microbiota dysbiosis | Postpartum dominance of pathogenic bacteria (e.g., Escherichia coli, Trueperella pyogenes) increases LPS burden and inflammatory tone | Sustains endometrial inflammation, disrupts mucosal immune tolerance, and inhibits uterine involution | Persistent systemic exposure to lipopolysaccharide and cytokines suppresses ovarian endocrine responsiveness | Reduced fertility and decreased pregnancy rates | [77, 78] |
AMPK = AMP-activated protein kinase, BHBA = β-hydroxybutyrate, CYP19A1 = Aromatase, FSH = Follicle-stimulating hormone, IL-1β = Interleukin-1 beta, IL-6 = Interleukin-6, LPS = Lipopolysaccharide, mTOR = Mechanistic target of rapamycin, NEFA = Non-esterified fatty acids, NF-κB = Nuclear factor kappa B, p38 MAPK = p38 mitogen-activated protein kinase, TLR4 = Toll-like receptor 4, TNF-α = Tumor necrosis factor-alpha.
UTERINE HEALTH AND ITS RELATIONSHIP WITH OVARIAN RECOVERY
Uterine health in dairy cows is tightly regulated by ovarian endocrine activity and represents a critical determinant of subsequent ovarian recovery and fertility outcomes [79]. Deficits in estrogen and progesterone directly alter uterine immune competence, myometrial contractility, and endometrial regeneration when ovarian function is impaired, particularly in cases of OH, thereby creating an intrauterine environment that further suppresses ovarian reactivation [80, 81]. Thus, OH and uterine dysfunction should be regarded as functionally interdependent rather than isolated disorders.
In postpartum dairy cows, uterine health is routinely assessed using a combination of vaginal discharge scoring, transrectal ultrasonography, and endometrial cytology, all of which provide clinically actionable information relevant to ovarian outcomes. Clinical examination focuses on discharge characteristics, whereas ultrasonography enables evaluation of uterine involution, luminal fluid accumulation, and endometrial thickness—parameters strongly associated with delayed resumption of ovarian cyclicity [82]. Endometrial cytology obtained via cytobrush or low-volume uterine lavage allows detection of subclinical endometritis based on polymorphonuclear neutrophil (PMN) proportions, a diagnostic approach particularly relevant in cows with OH that lack overt clinical signs.
Histopathologically, the uterus of cows with OH exhibits bovine-specific postpartum alterations, including reduced endometrial epithelial thickness, glandular epithelium degeneration, and persistent leukocyte infiltration within the lamina propria [83]. Unlike cyclical endometrial remodeling in nonpregnant cows, postpartum endometrial repair is highly dependent on timely estrogen and progesterone signaling. Estrogen deficiency leads to endometrial atrophy and reduced uterine gland secretory activity, compromising the uterine milieu required for sperm transport and early embryonic survival [84]. Concurrently, insufficient progesterone impairs stromal cell proliferation and downregulates growth factors such as insulin-like growth factor-1 (IGF-1) and transforming growth factor-β (TGF-β), both essential for postpartum uterine repair and subsequent luteal support [85].
Clinically, endometritis severity, classified as mild, moderate, or severe based on discharge characteristics and PMN thresholds, correlates closely with ovarian functional outcomes. Increasing uterine inflammatory severity is consistently associated with prolonged postpartum anestrus, reduced dominant follicle emergence, delayed first ovulation, and diminished luteal progesterone concentrations, indicating a direct relationship between uterine pathology and OH.
From an immunological standpoint, estrogen and progesterone play species-specific regulatory roles in the bovine uterine immune system [86]. Estrogen enhances uterine defense by stimulating antimicrobial peptides (e.g., β-defensin and lactoferrin) and promoting controlled recruitment of neutrophils and macrophages into the uterine lumen [87]. In contrast, progesterone limits excessive inflammation by suppressing proinflammatory cytokines such as IL-1β and TNF-α while promoting anti-inflammatory mediators including IL-10 [88]. In dairy cows with OH, prolonged deficiency of these hormones leads to immune dysregulation characterized by impaired phagocytic activity, reduced mucus secretion, and persistent endometrial inflammation, all of which compromise fertility under commercial production conditions [89, 90].
Delayed uterine involution is a frequent clinical finding in cows with OH during the postpartum period [91]. Under normal physiological conditions, uterine involution is largely completed within 30–45 days postpartum, and ovarian cyclicity resumes with the first ovulation typically occurring around 20–35 days postpartum [92, 93]. Deviations beyond these time frames serve as practical clinical indicators of pathological uterine dysfunction [94, 95]. Low estrogen and progesterone concentrations reduce myometrial contractility, impairing clearance of lochia and placental remnants [96], thereby increasing susceptibility to secondary bacterial infection and perpetuating ovarian suppression [97].
The relationship between uterine dysfunction and OH is reciprocal and self-reinforcing [98, 99]. Chronic uterine inflammation suppresses hypothalamic gonadotropin-releasing hormone (GnRH) secretion, reduces LH pulsatility, and inhibits follicular maturation [100]. Conversely, ovarian hormone deficiency further compromises uterine immune competence and tissue regeneration [101]. This feed-forward loop highlights the clinical importance of prioritizing uterine recovery as a prerequisite for restoring ovarian cyclicity and improving conception outcomes [102].
Figure 2 illustrates the proposed mechanistic link between uterine inflammation, hypothalamic–pituitary–ovarian axis disruption, and OH.
UTERINE FLUSHING THERAPY
Uterine flushing therapy is a non-hormonal intrauterine intervention primarily aimed at improving uterine health and facilitating postpartum uterine clearance in dairy cows, particularly in cases of mild to moderate endometritis and delayed uterine involution. Evidence-based studies consistently demonstrate improvements in uterine cleanliness and inflammatory status, whereas improvements in ovarian function should be interpreted as indirect secondary outcomes mediated through restoration of uterine–ovarian communication [103].
This approach involves intrauterine administration of sterile isotonic solutions, with or without adjunctive agents, to remove inflammatory exudate, reduce microbial load, and support endometrial repair [104].
Basic principles of flushing therapy
Under physiological postpartum conditions, uterine clearance is achieved through coordinated myometrial contractions and leukocyte-mediated phagocytosis [105]. Reduced estrogen and progesterone concentrations impair these mechanisms in cows with OH, leading to fluid retention and prolonged inflammation [106].
Multiple clinical studies report that intrauterine lavage using isotonic saline (0.9% NaCl) or Ringer’s lactate facilitates physical removal of lochia, necrotic debris, and inflammatory fluid, thereby accelerating uterine involution [107].
Although no universally standardized protocol exists, most field studies describe the use of 500–2000 mL of sterile isotonic solution per treatment, administered once or repeated up to two or three times at 24–72-h intervals depending on clinical severity. Flushing is generally performed after 21–28 days postpartum, when acute metritis has resolved [108]. Warmed solutions (38°C–40°C) have been associated with enhanced uterine contractility and improved endometrial perfusion.
Physiological and immunological effects
Beyond mechanical cleansing, uterine lavage increases endometrial microcirculation and stimulates myometrial activity, supporting faster involution [109]. Experimental and observational studies report enhanced leukocyte recruitment, particularly neutrophils and macrophages, following flushing [110].
At the mechanistic level, flushing therapy is proposed to modulate the local immune milieu, promoting a shift from persistent proinflammatory signaling toward resolution and tissue repair. Increased local expression of anti-inflammatory cytokines (e.g., IL-10 and TGF-β) has been reported, supporting epithelial regeneration [111].
Furthermore, reductions in intrauterine lipopolysaccharide (LPS) concentrations following lavage have been documented, which may attenuate LPS-mediated suppression of ovarian steroidogenesis via NF-κB signaling pathways [112, 113]. Although endocrine recovery is indirect, this provides a biologically plausible link between uterine flushing and gradual improvement in ovarian activity.
Adjunctive flushing agents: immunonutrition and phytotherapy
Immunonutritional or phytotherapeutic additives have been incorporated into flushing solutions [114]. Minerals such as calcium (Ca), zinc (Zn), and selenium (Se) support antioxidant defenses and innate immune function within uterine tissue [115]. Through its role in glutathione peroxidase activity, selenium supplementation has been associated with reduced oxidative damage and improved uterine recovery [116].
Phytotherapeutic compounds, including Aloe vera, Curcuma longa, and Azadirachta indica, have demonstrated antimicrobial and anti-inflammatory properties in experimental and field studies [117]. Bioactive molecules such as curcumin and aloin reduce TNF-α and IL-1β expression while promoting fibroblast proliferation and endometrial repair [118].
These adjuncts should be considered supportive rather than standalone treatments, and their efficacy is maximized when combined with appropriate herd nutrition and management [119].
Safety, contraindications, and procedural standardization
Uterine flushing should be performed using aseptic technique, sterile single-use catheters, and controlled infusion pressure. Potential risks include mechanical trauma, iatrogenic infection, and excessive uterine distension.
Flushing is contraindicated in cows with acute metritis, severe systemic illness, or confirmed pregnancy. Standardized biosecurity measures are essential to ensure animal welfare and reproducibility across clinical settings.
Comparison with hormonal therapy
Hormonal therapies (GnRH, PGF2α, or combinations) remain standard tools for managing OH [120]. Although these protocols are effective in inducing ovulation, they do not directly address uterine inflammation or metabolic imbalance [121].
In contrast, uterine flushing directly targets uterine pathology. Field studies report pregnancy rates of approximately 35%–55% in cows with mild to moderate endometritis following flushing therapy, which is comparable to or slightly higher than outcomes achieved with hormonal therapy alone [122].
Additionally, reductions in days open of approximately 10–25 days have been documented, particularly when flushing is integrated with nutritional and management interventions [10, 103, 123]. From an economic standpoint, flushing therapy generally involves lower direct costs and reduced reliance on repeated hormonal treatments, making it especially relevant for small- to medium-scale dairy systems.
Integrated and complementary use
Emerging evidence supports a sequential or combined strategy in which flushing therapy is applied to restore uterine health before hormonal synchronization. This approach improves the uterine environment prior to endocrine stimulation, thereby enhancing embryo survival [124].
Therefore, flushing therapy and hormonal protocols should be viewed as complementary, condition-based tools rather than competing interventions [125].
Implications for ovarian function and reproductive performance
Restoration of uterine health through flushing therapy reduces inflammatory and endotoxin-mediated suppression of the hypothalamic–pituitary–ovarian (HPO) axis, facilitating normalization of GnRH and LH secretion [126]. Improved endometrial perfusion and oxygenation further support steroidogenesis and follicular development [127].
Thus, uterine flushing represents a non-hormonal, immunomodulatory intervention with indirect yet clinically meaningful benefits for ovarian recovery and overall reproductive efficiency [128].
Immunometabolic perspectives in disease prevention and therapy
Reproductive health in dairy cows depends not only on hormonal balance but also on complex interactions between the immune system and metabolism, collectively termed immunometabolism [129]. In cases of OH, metabolic disturbances such as postpartum NEB play a major role in disrupting immune and endocrine homeostasis [130].
Immunometabolic strategies can be broadly categorized into (i) established nutritional and management-based practices currently applicable at the farm level and (ii) emerging or experimental interventions that require further validation. Decreased energy reserves and accumulation of metabolites such as NEFA and BHBA trigger oxidative stress and systemic inflammation, which in turn suppress ovarian function and worsen uterine conditions [131]. Accordingly, modern immunometabolic approaches emphasize feasible nutritional interventions and practical biomarker monitoring to prevent and manage reproductive disorders in a sustainable manner [132].
Immunometabolic strategies with high practical feasibility
In dairy cows, the transition period represents a critical phase during which energy and protein requirements increase to support early lactation while feed intake typically decreases [133]. Prolonged NEB reduces circulating glucose, insulin, and insulin-like growth factor-1 (IGF-1) concentrations, which directly inhibit ovarian follicular development [134]. Therefore, the most established and widely adopted immunometabolic strategy for preventing OH is optimization of energy balance through appropriate ration formulation and targeted supplementation [135].
Supplementation with omega-3 polyunsaturated fatty acids (PUFAs), particularly eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), is commonly administered at approximately 10–30 g/cow/day during the late dry period through early lactation. These fatty acids exert systemic anti-inflammatory effects by suppressing prostaglandin E2 (PGE2) and leukotriene B2 (LTB2) synthesis, thereby reducing uterine inflammation and supporting endometrial recovery [136].
Trace mineral supplementation, including selenium (0.3 mg/kg DM), zinc (40–60 mg/kg DM), and copper (10–15 mg/kg DM), is routinely implemented in commercial dairy systems and supports antioxidant defense mechanisms essential for ovarian and uterine recovery [137]. Similarly, vitamin E (1,000–3,000 IU/cow/day) and β-carotene (300–500 mg/cow/day) supplementation during the transition period has demonstrated consistent benefits under field conditions, including improved leukocyte function and faster resolution of postpartum uterine inflammation [138, 139].
Emerging and experimental immunometabolic approaches
Several immunometabolic interventions remain experimental or are primarily supported by controlled or pilot studies. These include the use of specific cytokine biomarkers (e.g., IL-6 and IL-8), nutraceutical antioxidants, and advanced delivery systems such as nano-formulated supplements [140]. Although these approaches provide valuable mechanistic insights, their routine application is currently limited by cost, assay availability, and regulatory considerations [141].
Compounds such as resveratrol, curcumin, and N-acetylcysteine have shown potential to modulate NF-κB and mTOR signaling pathways and improve insulin sensitivity in reproductive tissues [142]. However, these compounds should be regarded as adjunctive or investigational until validated through large-scale field trials.
Practical feasibility and on-farm implementation
Successful adoption of immunometabolic strategies depends on feasibility, cost-effectiveness, and ease of integration into existing herd management programs [143]. Although rumen-protected supplements offer improved bioavailability, their higher cost may limit their use in small- to medium-scale dairy operations [144, 145]. In contrast, monitoring metabolic indicators such as BHBA and NEFA using handheld devices is highly feasible at the farm level and allows early identification of cows at risk of OH [146].
Therefore, a tiered implementation strategy is recommended, prioritizing readily measurable metabolic indicators and established nutritional interventions while reserving laboratory-based biomarkers and experimental nutraceuticals for high-risk cows or research-focused herds.
Integrated preventive framework
A multidisciplinary strategy encompassing nutritional management, uterine infection control, and supportive non-hormonal therapies is required for an effective immunometabolic-based preventive and therapeutic approach [103]. By prioritizing established, feasible interventions and selectively incorporating emerging approaches, immunometabolic management can realistically improve reproductive efficiency while minimizing reliance on hormonal treatments [147].
Biomarker-based monitoring in immunometabolic reproductive management
This subsection focuses specifically on biomarker-based monitoring and its practical application to avoid conceptual overlap with earlier sections, rather than reiterating general immunometabolic mechanisms. Biomarker monitoring provides an objective framework for identifying cows at risk of OH and associated uterine disorders, particularly during the transition and early postpartum periods [148].
For editorial clarity and field applicability, immunometabolic biomarkers can be categorized into three functional groups: metabolic, inflammatory, and endocrine biomarkers, each serving a distinct diagnostic purpose.
Metabolic biomarkers
Metabolic parameters are the most practical and widely used indicators for on-farm reproductive risk monitoring. Circulating concentrations of NEFA, BHBA, glucose, and IGF-1 reflect energy balance and metabolic stress during the transition period [149].
Plasma NEFA concentrations exceeding approximately 0.3–0.4 mmol/L during the late prepartum period and >0.6–0.7 mmol/L during the early postpartum period are commonly associated with an increased risk of OH and uterine disease [150]. Similarly, postpartum BHBA concentrations above 1.2–1.4 mmol/L indicate subclinical ketosis and impaired reproductive performance. Reduced IGF-1 concentrations during the first 2–4 weeks postpartum have been consistently linked to delayed ovulation and reduced dominant follicle growth [151].
From a practical standpoint, BHBA and, to a lesser extent, NEFA can be measured using handheld or semi-automated analyzers, making metabolic biomarkers highly feasible for routine herd-level screening.
Inflammatory biomarkers
Inflammatory biomarkers provide insight into systemic and uterine immune activation but are generally better suited for targeted or confirmatory diagnostics than routine screening. Acute-phase proteins such as haptoglobin, ceruloplasmin, and albumin reflect the balance between inflammatory processes and nutritional status [152].
Haptoglobin concentrations below approximately 0.1 g/L are typically considered indicative of normal postpartum adaptation, whereas values exceeding 0.3–0.5 g/L during early lactation are frequently associated with subclinical inflammation, endometritis, and impaired luteal function. Elevated ceruloplasmin concentrations are likewise associated with uterine inflammatory disorders [153].
Cytokines such as IL-6 and IL-8 play key roles in local uterine immune responses. Although cytokine measurement is mainly confined to research settings, persistently elevated IL-8 concentrations in uterine fluid or serum may indicate sustained neutrophil recruitment and an unfavorable uterine environment for embryo implantation [154].
Endocrine biomarkers
Endocrine biomarkers link metabolic and inflammatory status to ovarian function. Hormones such as IGF-1, insulin, progesterone, and estradiol provide complementary information on hypothalamic–pituitary–ovarian (HPO) axis activity [155].
Reduced IGF-1 and insulin concentrations during early lactation are associated with delayed follicular development and reflect prolonged NEB. Low circulating progesterone concentrations during the expected luteal phase may indicate impaired luteal function secondary to metabolic or inflammatory stress rather than primary ovarian failure [156].
Temporal application and monitoring strategies
The diagnostic value of biomarkers is maximized when measurements are aligned with key physiological windows, including late prepartum (−21 to −7 days), early postpartum (0–14 days), and the peak reproductive risk period (14–35 days postpartum). Repeated measurements across these periods provide greater predictive accuracy than single-time-point sampling [157].
A tiered monitoring strategy is recommended to enhance practical applicability:(i) routine on-farm screening using metabolic biomarkers (BHBA and NEFA) for all transition cows, and(ii) targeted laboratory-based inflammatory or endocrine testing in cows identified as high-risk based on metabolic profiles or clinical findings [158].
Integration into reproductive decision-making
When integrated with nutritional management and uterine health assessment, biomarker-based monitoring enables earlier and more targeted interventions, including nutritional adjustment, uterine flushing, or supportive non-hormonal therapies [149, 150]. This approach allows prevention or mitigation of OH before irreversible reproductive dysfunction occurs, thereby improving reproductive efficiency while reducing reliance on blanket hormonal treatments.
FUTURE RESEARCH DIRECTIONS
Based on the key gaps identified in earlier sections, including limited field validation of immunometabolic biomarkers, lack of standardized non-hormonal intervention protocols, and challenges in translating experimental findings into herd-level practice, future research in immunometabolic-based reproductive management can be systematically organized into short-, medium-, and long-term priorities. This structured framework aims to facilitate progressive development from mechanistic understanding to practical implementation.
Short-term research priorities: mechanistic validation and feasibility studies
In the short-term, research should focus on addressing the current lack of validated non-hormonal alternatives and biomarker-guided interventions identified in this review. Priority should be given to proof-of-concept and validation studies evaluating non-hormonal metabolic modulators capable of restoring immune–metabolic balance during the transition period [159].
Compounds such as resveratrol, curcumin, and N-acetylcysteine have demonstrated antioxidant and metabolic regulatory potential, including suppression of NF-κB and mTOR signaling pathways and improvement of insulin sensitivity and mitochondrial function [160]. However, their efficacy has largely been demonstrated under experimental or controlled conditions. Therefore, short-term studies should focus on dose optimization, safety evaluation, and biomarker-based assessment of reproductive responses, directly addressing the current gap in standardized efficacy metrics.
Medium-term research priorities: translational optimization and protocol integration
Medium-term research should aim to bridge the translational gap between experimental findings and applied reproductive management. This includes optimization studies evaluating delivery systems, formulation stability, and interactions with conventional nutritional strategies [161].
The development of nanotechnology-based immunonutrition formulations represents a promising approach to overcome bioavailability limitations and inconsistent absorption highlighted earlier in the manuscript [162]. At this stage, uterine flushing therapy, immunometabolic supplementation, and precision biomarker monitoring should be integrated into standardized, condition-based reproductive protocols, allowing adaptive responses to metabolic stress and uterine inflammation.
Long-term research priorities: system-level implementation and sustainability
Long-term research should address the current lack of large-scale, system-level evidence supporting immunometabolic reproductive strategies. Applied studies should focus on identifying specific molecular targets linking metabolic stress to ovarian and endometrial dysfunction and validating intervention strategies across diverse production systems [163].
Multicenter field trials evaluating reproductive performance, animal welfare, economic feasibility, and environmental sustainability should complement these efforts. Such studies are essential to confirm that immunometabolic approaches can achieve durable fertility improvements while reducing reliance on hormonal treatments, thereby supporting sustainable dairy production [164].
Regulatory, ethical, and adoption considerations
In parallel with biological research, regulatory and adoption-related challenges identified earlier in the manuscript must be systematically addressed. Comprehensive toxicological and residue studies are required to ensure the long-term safety of non-hormonal metabolic modulators, particularly nano-formulated compounds, and to ensure compliance with food safety and animal welfare standards [165].
Regulatory approval pathways for novel feed additives and advanced formulations may vary across regions, potentially limiting widespread adoption. Additionally, practical constraints such as cost, ease of application, farmer awareness, and integration into existing herd management programs represent critical barriers to on-farm implementation [166, 167]. Addressing these challenges through interdisciplinary collaboration, stakeholder engagement, and extension-focused research will be essential for successful translation of immunometabolic innovations into routine dairy reproductive management [168].
Figure 3 summarizes the proposed mechanistic framework linking postpartum uterine flushing, immunometabolic regulation, and restoration of ovarian function.
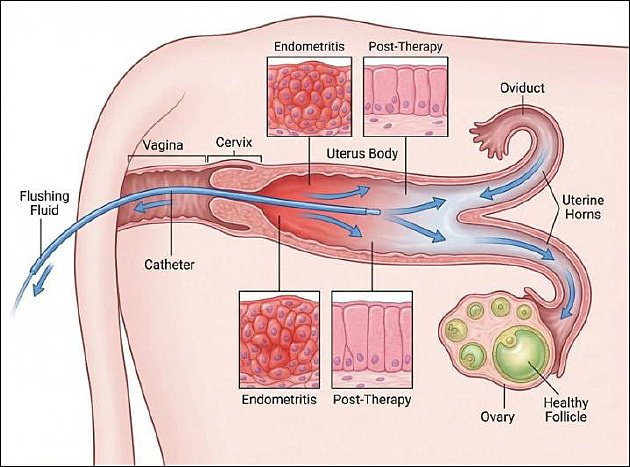
Figure 3. Intrauterine flushing therapy and ovarian recovery in dairy cows. This schematic illustration was generated using artificial intelligence tools (ChatGPT 5.2) and subsequently modified by the authors.
CONCLUSION
This review highlights that OH in dairy cows is not an isolated ovarian disorder but a multifactorial condition arising from interconnected endocrine, metabolic, immune, and uterine processes. Evidence synthesized across physiological, clinical, and immunometabolic studies demonstrates that postpartum NEB, oxidative stress, and uterine inflammation act synergistically to disrupt hypothalamic–pituitary–ovarian signaling, impair follicular development, and delay the resumption of ovarian cyclicity. The bidirectional relationship between uterine health and ovarian function underscores the importance of considering reproductive disorders within an integrated immunometabolic framework rather than as separate pathological entities.
From a practical standpoint, this integrative perspective supports a shift toward mechanism-based reproductive management. Biomarker-guided monitoring of metabolic indicators such as NEFA, BHBA, and IGF-1 enables early identification of cows at risk, while nutritional optimization and transition period management remain foundational preventive strategies. Uterine flushing therapy emerges as a physiologically oriented, non-hormonal intervention that promotes uterine clearance, immune resolution, and endometrial repair, with indirect yet clinically meaningful benefits for ovarian recovery and fertility outcomes. When applied in combination with targeted metabolic support and reproductive monitoring, such approaches may reduce reliance on blanket hormonal treatments and improve herd-level reproductive efficiency.
A key strength of this review lies in its comprehensive integration of endocrine, metabolic, immune, and clinical evidence into a unified conceptual model linking uterine inflammation, immunometabolic imbalance, and ovarian dysfunction. However, limitations include reliance on heterogeneous experimental and field studies, variable biomarker thresholds across production systems, and limited large-scale validation of emerging non-hormonal interventions.
Future research should prioritize multicenter field trials, standardized biomarker-based protocols, and translational studies evaluating integrated management strategies under diverse dairy production conditions. Ultimately, adopting an immunometabolic-based approach that combines early risk detection, uterine health restoration, and supportive nutritional management offers a promising pathway to enhance reproductive performance, animal welfare, and sustainability in modern dairy production systems.
Overall, the evidence supports immunometabolic-based reproductive management as a realistic pathway to improve postpartum uterine recovery and accelerate ovarian reactivation while enhancing animal welfare and reducing dependence on repeated hormonal treatments. Implementing feasible on-farm screening (e.g., BHBA and NEFA), strengthening transition cow nutrition, and applying uterine-focused non-hormonal therapies in appropriately selected cows can collectively deliver more sustainable fertility outcomes in modern dairy production systems.
DATA AVAILABILITY
All the generated data are included in the manuscript. The selection of articles and supplementary text can be made available from the corresponding author upon a request.
AUTHOR’S CONTRIBUTIONS
CB, IM, and ARK: Drafted the manuscript. SM, TWS, and ES: Revised and edited the manuscript. SR, RZA, and LP: Participated in preparation and critically reviewed the manuscript. BPP, WW, and SS: Edited the references. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
This work was supported by the Education Fund Management Institution (LPDP) and the Higher Education Financing Center (BPPT) under the Indonesian Education Scholarship Program (BPI), Ministry of Education, Culture, Research, and Technology (MoECRT), Indonesia. Grant number: 01366/BPPT/BPI.06/9/2023.
REFERENCES
- Tolosa F, Netsere M, Habtamu Y. Assessment of Major Reproductive Disorders in Dairy Cattle in and around Bale Robe, Oromia Regional State, Ethiopia. Vet Med Int 2021;2021(1):8855718. [Google Scholar] | [Crossref]
- Villar SL, Pérez-Marín CC, Álvarez J, Acción A, Barrionuevo R, Becerra JJ, Peña AI, Herradón PG, Quintela LA, Yáñez U. A Prospective Study of the Prevalence and Predictive Risk Factors of Repeat Breeder Syndrome in Dairy Cattle in the North of Spain. Animals (Basel) 2025;15(2):266. [Google Scholar] | [Crossref]
- Carbonari A, Burgio M, Frattina L, Cicirelli V, Rizzo A. Repeat Breeder Syndrome Therapies in Dairy Cows:A Review. Reprod Domest Anim 2024;59(10):e14732. [Google Scholar] | [Crossref]
- Triwutanon S, Rukkwamsuk T. Factors affecting first ovulation in postpartum dairy cows under tropical conditions:A review. Open Vet J 2023;13(12):1536-1542. [Google Scholar] | [Crossref]
- Khamitova L, Rudakov R, Knyazeva M, Metlyakova A. Hormonal therapy for ovarian dysfunctions in high-productive cows. BIO Web Conf 2020;17(1):00205. [Google Scholar] | [Crossref]
- Wang F, Guo Y, Su X, Cao J. Choline as a Modulator of Periparturient Diseases in Dairy Cows. Vet Sci 2025;12(10):1016. [Google Scholar] | [Crossref]
- Chaudhary V, Haloi P, Munjal K, Chitme HR. Pharmacotherapeutic importance of phytoconstituents in the management of Polycystic Ovary Syndrome (PCOS) and associated complications. Egypt J Basic Appl Sci 2025;12(1):1-26. [Google Scholar] | [Crossref]
- Šišljagić D, Blažetić S, Heffer M, Delać MV, Muller A. The Interplay of Uterine Health and Obesity:A Comprehensive Review. Biomedicines 2024;12(12):2801. [Google Scholar] | [Crossref]
- Ramil UY, Jezierska S, Krupa M, Pascottini OB. Fundamentals of microbiome-based therapies for reproductive tract inflammatory diseases in domestic animals. Anim Reprod 2025;22(3):e20250030. [Google Scholar] | [Crossref]
- Hermadi HA, Khairullah AR, Damayanti Y, Safitri E, Tyasningsih W, Warsito SH, Kusala MKJ, Wibowo S, Moses IB, Kurniawan MA, Wardhani BWK, Prasetyo ST, Kurniasih DAA, Rahajeng ADR, Fauziah I, Vidhianty CB, Ansori ANM, Wasito W, Ahmad RZ. Endometritis in Cattle:A Review of Current Understanding and Practical Causes of Repeat Breeding. Trends Sci 2025;22(6):9587. [Google Scholar] | [Crossref]
- Datkhayeva Z, Iskakova A, Mireeva A, Seitaliyeva A, Skakova R, Kulniyazova G, Shayakhmetova A, Koshkimbayeva G, Sarmuldayeva C, Nurseitova L, Koshenova L, Imanbekova G, Maxutova D, Yerkenova S, Shukirbayeva A, Pernebekova U, Dushimova Z, Amirkhanova A. The Multifactorial Pathogenesis of Endometriosis:A Narrative Review Integrating Hormonal, Immune, and Microbiome Aspects. Medicina (Kaunas) 2025;61(5):811. [Google Scholar] | [Crossref]
- Xu R, He X, Xu J, Yu G, Wu Y. Immunometabolism:signaling pathways, homeostasis, and therapeutic targets. MedComm (2020) 2024;5(11):e789. [Google Scholar] | [Crossref]
- Park W, Lim W, Kim M, Jang H, Park SJ, Song G, Park S. Female reproductive disease, endometriosis:From inflammation to infertility. Mol Cells 2025;48(1):100164. [Google Scholar] | [Crossref]
- Parisi F, Milazzo R, Savasi VM, Cetin I. Maternal Low-Grade Chronic Inflammation and Intrauterine Programming of Health and Disease. Int J Mol Sci 2021;22(4):1732. [Google Scholar] | [Crossref]
- Howard SA, Benhabbour SR. Non-Hormonal Contraception. J Clin Med 2023;12(14):4791. [Google Scholar] | [Crossref]
- Hosseini E, Aghajanpour S, Chekini Z, Zameni N, Zolfaghary Z, Aflatoonian R, Hafezi M. An Approach to Improve Endometrial Receptivity:Is It Beneficial to Flush The Uterine Cavity with Follicular Fluid and Granulosa Cells?A Phase III Randomised Clinical Trial. Int J Fertil Steril 2024;18((Suppl 1)):22-29. [Google Scholar] | [Crossref]
- Mikhael S, Punjala-Patel A, Gavrilova-Jordan L. Hypothalamic–Pituitary–Ovarian Axis Disorders Impacting Female Fertility. Biomedicines 2019;7(1):5. [Google Scholar] | [Crossref]
- Kasimanickam R, Kasimanickam V, Ferreira J, Kastelic J, de Souza F. Regulatory RNA Networks in Ovarian Follicular Cysts in Dairy Cows:Implications for Human Polycystic Ovary Syndrome. Genes 2025;16((7)):791. [Google Scholar] | [Crossref]
- Xu XL, Huang ZY, Yu K, Li J, Fu XW, Deng SL. Estrogen Biosynthesis and Signal Transduction in Ovarian Disease. Front Endocrinol (Lausanne) 2022;13(1):827032. [Google Scholar] | [Crossref]
- Ajayi AF, Akhigbe RE. Staging of the estrous cycle and induction of estrus in experimental rodents:an update. Fertil Res Pract 2020;6(1):5. [Google Scholar] | [Crossref]
- Coss D. Regulation of reproduction via tight control of gonadotropin hormone levels. Mol Cell Endocrinol 2018;463(1):116-130. [Google Scholar] | [Crossref]
- Morton AJ, Candelaria JI, McDonnell SP, Zgodzay DP, Denicol AC. Review:Roles of follicle-stimulating hormone in preantral folliculogenesis of domestic animals:what can we learn from model species and where do we go from here?. Animal 2023;17((Suppl 1)):100743. [Google Scholar] | [Crossref]
- Pan Y, Pan C, Zhang C. Unraveling the complexity of follicular fluid:insights into its composition, function, and clinical implications. J Ovarian Res 2024;17((1)):237. [Google Scholar] | [Crossref]
- Chen YH, Chu TY. Ovulation:A consequence of acute inflammation cultivated by E2-induced reactive oxygen species and triggered by progesterone withdrawal. Tzu Chi Med J 2025;37(4):351-359. [Google Scholar] | [Crossref]
- Bulletti C, Bulletti FM, Sciorio R, Guido M. Progesterone:The Key Factor of the Beginning of Life. Int J Mol Sci 2022;23(22):14138. [Google Scholar] | [Crossref]
- Phong V, Fujikawa T, Kawano K, Yanagawa Y, Katagiri S. Effects of prostaglandin F2αtreatment at follicular wave emergence on endometrial epidermal growth factor concentration on day 3 of the next estrous cycle and on fertility in dairy cows. Theriogenology 2023;212(1):189-196. [Google Scholar] | [Crossref]
- Sammad A, Khan MZ, Abbas Z, Hu L, Ullah Q, Wang Y, Zhu H, Wang Y. Major Nutritional Metabolic Alterations Influencing the Reproductive System of Postpartum Dairy Cows. Metabolites 2022;12(1):60. [Google Scholar] | [Crossref]
- Walsh S, Delaby L, Kennedy M, Galvin N, McKay ZC, Egan M. Intake profile, milk production, and energy balance of early lactation spring-calving Holstein Friesian and Jersey ×Holstein Friesian dairy cows in high-utilization pasture-based systems. J Dairy Sci 2024;107(10):8058-8071. [Google Scholar] | [Crossref]
- Fenwick MA, Llewellyn S, Fitzpatrick R, Kenny DA, Murphy JJ, Patton J, Wathes DC. Negative energy balance in dairy cows is associated with specific changes in IGF-binding protein expression in the oviduct. Reproduction 2008;135(1):63-75. [Google Scholar] | [Crossref]
- Blüher S, Shah S, Mantzoros CS. Leptin deficiency:clinical implications and opportunities for therapeutic interventions. J Investig Med 2009;57(7):784-788. [Google Scholar] | [Crossref]
- Athar F, Karmani M, Templeman NM. Metabolic hormones are integral regulators of female reproductive health and function. Biosci Rep 2024;44(1):BSR20231916. [Google Scholar] | [Crossref]
- Huang Y, Liu Z, Geng Y, Li F, Hu R, Song Y, Zhang M, Song K. The risk factors, pathogenesis and treatment of premature ovarian insufficiency. J Ovarian Res 2025;18(1):134. [Google Scholar] | [Crossref]
- Juszczyk G, Mikulska J, Kasperek K, Pietrzak D, Mrozek W, Herbet M. Chronic Stress and Oxidative Stress as Common Factors of the Pathogenesis of Depression and Alzheimer's Disease:The Role of Antioxidants in Prevention and Treatment. Antioxidants (Basel) 2021;10(9):1439. [Google Scholar] | [Crossref]
- Liu H, Jin L, Wang X, Shi J, He Y, Sun N, Yang F. Reactive oxygen species in polycystic ovary syndrome:Mechanistic insights into pathogenesis and therapeutic opportunities. Redox Biol 2025;85(1):103776. [Google Scholar] | [Crossref]
- Wang L, Xiong D, Yan H, Long F, Zhang G, Zeng J, Liu J, Zeng Q, Zhang Y, Bai X, Luo M, Liu W. MMP-9 dysregulation and chronic inflammation in polycystic ovary syndrome:linking ovulatory dysfunction to diagnostic implications. J Ovarian Res 2025;18(1):247. [Google Scholar] | [Crossref]
- Lonardo MS, Cacciapuoti N, Guida B, Di Lorenzo M, Chiurazzi M, Damiano S, Menale C. Hypothalamic-Ovarian axis and Adiposity Relationship in Polycystic Ovary Syndrome:Physiopathology and Therapeutic Options for the Management of Metabolic and Inflammatory Aspects. Curr Obes Rep 2024;13(1):51-70. [Google Scholar] | [Crossref]
- Bruinjé TC, LeBlanc SJ. Graduate Student Literature Review:Implications of transition cow health for reproductive function and targeted reproductive management. J Dairy Sci 2024;107(10):8234-8246. [Google Scholar] | [Crossref]
- Salman A, Prihatno SA, Sumiarto B. epidemiological analysis of ovarium hypofunction in beef cattle in Jepara regency. Vet Pract 2020;21(2):274-278. [Google Scholar] | [Crossref]
- Wang R, Lv Y, Dou T, Yang Q, Yu C, Guan Q. Autoimmune thyroid disease and OH:a review of literature. J Ovarian Res 2024;17(1):125. [Google Scholar] | [Crossref]
- Howard SR. Interpretation of reproductive hormones before, during and after the pubertal transition—Identifying health and disordered puberty. Clin Endocrinol (Oxf) 2021;95(5):702-715. [Google Scholar] | [Crossref]
- Smedlund KB, Hill JW. The role of non-neuronal cells in hypogonadotropic hypogonadism. Mol Cell Endocrinol 2020;518(1):110996. [Google Scholar] | [Crossref]
- Recchia K, Jorge AS, Pessôa LVF, Botigelli RC, Zugaib VC, de Souza AF, Martins DDS, Ambrósio CE, Bressan FF, Pieri NCG. Actions and Roles of FSH in Germinative Cells. Int J Mol Sci 2021;22(18):10110. [Google Scholar] | [Crossref]
- Laksmi DNDI, Trilaksana IGNB, Sukernayasa IW, Gunawan IWNF, Merdana IM. The efficacy of bovine pituitary extract for treating ovarian hypofunction cases in Bali cattle. Int J Vet Sci 2024;13(5):687-690. [Google Scholar] | [Crossref]
- Zhang T, Lin M, Wang C, Zhou J. Mechanisms of follicular atresia:focus on apoptosis, autophagy, and ferroptosis. Front Endocrinol (Lausanne) 2025;16(1):1603467. [Google Scholar] | [Crossref]
- Przygrodzka E, Plewes MR, Davis JS. Luteinizing Hormone Regulation of Inter-Organelle Communication and Fate of the Corpus Luteum. Int J Mol Sci 2021;22(18):9972. [Google Scholar] | [Crossref]
- Wang J, Zhan H, Wang Y, Zhao L, Huang Y, Wu R. Current advances in understanding endometrial epithelial cell biology and therapeutic applications for intrauterine adhesion. Stem Cell Res Ther 2024;15(1):379. [Google Scholar] | [Crossref]
- Ahmad F, Ahmed SH, Choucair F, Chouliaras S, Awwad J, Terranegra A. A disturbed communication between hypothalamic-pituitary-ovary axis and gut microbiota in female infertility:is diet to blame?. J Transl Med 2025;23(1):92. [Google Scholar] | [Crossref]
- Qiao K, Jiang R, Contreras GA, Xie L, Pascottini OB, Opsomer G, Dong Q. The Complex Interplay of Insulin Resistance and Metabolic Inflammation in Transition Dairy Cows. Animals (Basel) 2024;14(6):832. [Google Scholar] | [Crossref]
- Lőrincz CE, Börzsei D, Hoffmann A, Varga C, Szabó R. Mechanisms and Target Parameters in Relation to Polycystic Ovary Syndrome and Physical Exercise:Focus on the Master Triad of Hormonal Changes, Oxidative Stress, and Inflammation. Biomedicines 2024;12(3):560. [Google Scholar] | [Crossref]
- Odle AK, Akhter N, Syed MM, Allensworth-James ML, Beneš H, Castillo AIM, MacNicol MC, MacNicol AM, Childs GV. Leptin Regulation of Gonadotrope Gonadotropin-Releasing Hormone Receptors As a Metabolic Checkpoint and Gateway to Reproductive Competence. Front Endocrinol (Lausanne) 2018;8(1):367. [Google Scholar] | [Crossref]
- Strickland JM, Wisnieski L, Mavangira V, Sordillo LM. Serum Vitamin D Is Associated with Antioxidant Potential in Peri-Parturient Cows. Antioxidants (Basel) 2021;10(9):1420. [Google Scholar] | [Crossref]
- Yan F, Zhao Q, Li Y, Zheng Z, Kong X, Shu C, Liu Y, Shi Y. The role of oxidative stress in ovarian aging:a review. J Ovarian Res 2022;15(1):100. [Google Scholar] | [Crossref]
- Brinca AT, Peiró AM, Evangelio PM, Eleno I, Oliani AH, Silva V, Vicente LF, Ramalhinho AC, Gallardo E. Follicular Fluid and Blood Monitorization of Infertility Biomarkers in Women with Endometriosis. Int J Mol Sci 2024;25(13):7177. [Google Scholar] | [Crossref]
- Liu T, Huang Y, Lin H. Estrogen disorders:Interpreting the abnormal regulation of aromatase in granulosa cells (Review). Int J Mol Med 2021;47(5):73. [Google Scholar] | [Crossref]
- Shahzad M, Cao J, Kolachi HA, Ayantoye JO, Yu Z, Niu Y, Wan P, Zhao X. Unravelling the Signature Follicular Fluid Metabolites in Dairy Cattle Follicles Growing Under Negative Energy Balance:An In Vitro Approach. Int J Mol Sci 2024;25(23):12629. [Google Scholar] | [Crossref]
- Bionaz M, Vargas-Bello-Pérez E, Busato S. Advances in fatty acids nutrition in dairy cows:from gut to cells and effects on performance. J Anim Sci Biotechnol 2020;11(1):110. [Google Scholar] | [Crossref]
- Lei Z, Ali I, Yang M, Yang C, Li Y, Li L. Non-Esterified Fatty Acid-Induced Apoptosis in Bovine Granulosa Cells via ROS-Activated PI3K/AKT/FoxO1 Pathway. Antioxidants (Basel) 2023;12(2):434. [Google Scholar] | [Crossref]
- Raval FM, Nikolajczyk BS. The Bidirectional Relationship between Metabolism and Immune Responses. Discoveries (Craiova) 2013;1(1):e6. [Google Scholar] | [Crossref]
- Meikle A, de Brun V, Carriquiry M, Soca P, Sosa C, Adrien ML, Chilibroste P, Abecia JA. Influences of nutrition and metabolism on reproduction of the female ruminant. Anim Reprod 2018;15((Suppl 1)):899-911. [Google Scholar] | [Crossref]
- Bruinjé TC, LeBlanc SJ. Invited Review:Inflammation and Health in the Transition Period Influence Reproductive Function in Dairy Cows. Animals (Basel) 2025;15(5):633. [Google Scholar] | [Crossref]
- Antanaitis R, Džermeikaitė K, Januškevičius V, Šimonytė I, Baumgartner W. In-Line Registered Milk Fat-to-Protein Ratio for the Assessment of Metabolic Status in Dairy Cows. Animals (Basel) 2023;13(20):3293. [Google Scholar] | [Crossref]
- Khan MZ, Huang B, Kou X, Chen Y, Liang H, Ullah Q, Khan IM, Khan A, Chai W, Wang C. Enhancing bovine immune, antioxidant and anti-inflammatory responses with vitamins, rumen-protected amino acids, and trace minerals to prevent periparturient mastitis. Front Immunol 2024;14(1):1290044. [Google Scholar] | [Crossref]
- Li C, Huang J, Chen X, Yan Y, Li L, Zhao W. Transcriptome Analysis Reveals That NEFA and β-Hydroxybutyrate Induce Oxidative Stress and Inflammatory Response in Bovine Mammary Epithelial Cells. Metabolites 2022;12(11):1060. [Google Scholar] | [Crossref]
- Molnar V, Matišić V, Kodvanj I, Bjelica R, Jeleč Ž, Hudetz D, Rod E, Čukelj F, Vrdoljak T, Vidović D, Starešinić M, Sabalić S, Dobričić B, Petrović T, Antičević D, Borić I, Košir R, Zmrzljak UP, Primorac D. Cytokines and Chemokines Involved in Osteoarthritis Pathogenesis. Int J Mol Sci 2021;22(17):9208. [Google Scholar] | [Crossref]
- Adamczak R, Ukleja-Sokołowska N, Lis K, Dubiel M. Function of Follicular Cytokines:Roles Played during Maturation, Development and Implantation of Embryo. Medicina (Kaunas) 2021;57(11):1251. [Google Scholar] | [Crossref]
- Tobolski D, Krupa M, Polak Z, Pascottini OB, Opsomer G, Barański W. Exploring the influence of endometritis diagnostic criteria on uterine involution, milk yield and fertility in dairy cows. BMC Vet Res 2025;21(1):276. [Google Scholar] | [Crossref]
- Khan KN, Fujishita A, Hiraki K, Kitajima M, Nakashima M, Fushiki S, Kitawaki J. Bacterial contamination hypothesis:a new concept in endometriosis. Reprod Med Biol 2018;17(2):125-133. [Google Scholar] | [Crossref]
- Wang K, Lai W, Min T, Wei J, Bai Y, Cao H, Guo J, Su Z. The Effect of Enteric-Derived Lipopolysaccharides on Obesity. Int J Mol Sci 2024;25(8):4305. [Google Scholar] | [Crossref]
- Guijarro-Muñoz I, Compte M, Álvarez-Cienfuegos A, Álvarez-Vallina L, Sanz L. Lipopolysaccharide activates Toll-like receptor 4 (TLR4)-mediated NF-κB signaling pathway and proinflammatory response in human pericytes. J Biol Chem 2014;289(4):2457-2468. [Google Scholar] | [Crossref]
- Magata F, Kikuzawa M, Bollwein H, Matsuda F, Haneda S. Lipopolysaccharide-binding protein in follicular fluid is associated with the follicular inflammatory status and granulosa cell steroidogenesis in dairy cows. J Reprod Dev 2024;70(3):169-176. [Google Scholar] | [Crossref]
- Garza-Lombó C, Schroder A, Reyes-Reyes EM, Franco R. mTOR/AMPK signaling in the brain:Cell metabolism, proteostasis and survival. Curr Opin Toxicol 2018;8(1):102-110. [Google Scholar] | [Crossref]
- Alasadi A, Fadhil N, Chen S. Deciphering the critical roles of the AMPK/mTOR signaling pathway in cancer cell metabolism (Review). World Acad Sci J 2025;7(1):103. [Google Scholar] | [Crossref]
- Qi Q, Li Y, Chen Z, Luo Z, Zhou T, Zhou J, Zhang Y, Chen S, Wang L. Update on the pathogenesis of endometriosis-related infertility based on contemporary evidence. Front Endocrinol (Lausanne) 2025;16(1):1558271. [Google Scholar] | [Crossref]
- Sheldon IM, Cronin JG, Pospiech M, Turner ML. Symposium review:Mechanisms linking metabolic stress with innate immunity in the endometrium. J Dairy Sci 2018;101(4):3655-3664. [Google Scholar] | [Crossref]
- Blazheva S, Pachkova S, Bodurska T, Ivanov P, Blazhev A, Lukanov T, Konova E. Unlocking the Uterine Code:Microbiota, Immune Cells, and Therapy for Recurrent Reproductive Failure. Microorganisms 2024;12(3):547. [Google Scholar] | [Crossref]
- Mora PE, Valbuena D, Diez-Juan A. The Role of the Gut Microbiota in Female Reproductive and Gynecological Health:Insights into Endometrial Signaling Pathways. Life (Basel) 2025;15(5):762. [Google Scholar] | [Crossref]
- LeBlanc SJ. Review:Postpartum reproductive disease and fertility in dairy cows. Animal 2023;17((Suppl 1)):100781. [Google Scholar] | [Crossref]
- LeBlanc SJ. Postpartum uterine disease and dairy herd reproductive performance:a review. Vet J 2008;176(1):102-114. [Google Scholar] | [Crossref]
- Matsuno Y, Imakawa K. Biological Aging and Uterine Fibrosis in Cattle:Reproductive Trade-Offs from Enhanced Productivity. Cells 2025;14(13):955. [Google Scholar] | [Crossref]
- Seekford ZK, Tariq A, Macay GA, Jenkins KM, Dickson MJ, Melo GD, Pohler KG, Sheldon IM, Bromfield JJ. Uterine disease in dairy cows is associated with contemporaneous perturbations to ovarian function. Theriogenology 2025;232(1):20-29. [Google Scholar] | [Crossref]
- Klonos E, Katopodis P, Karteris E, Papanikolaou E, Tarlatzis B, Pados G. Endometrial changes in estrogen and progesterone receptor expression during implantation in an oocyte donation program. Exp Ther Med 2020;20(6):178. [Google Scholar] | [Crossref]
- Várhidi Z, Csikó G, Bajcsy ÁC, Jurkovich V. Uterine Disease in Dairy Cows:A Comprehensive Review Highlighting New Research Areas. Vet Sci 2024;11(2):66. [Google Scholar] | [Crossref]
- Thasmi CN, Siregar TN, Wahyuni S, Aliza D, Panjaitan B, Nazaruddin N, Sabila FN, Fallatanza M. Anatomical and histological changes of uterine horn of Aceh cattle with repeat breeding. J Adv Vet Anim Res 2018;5(4):445-453. [Google Scholar] | [Crossref]
- Bonavina G, Taylor HS. Endometriosis-associated infertility:From pathophysiology to tailored treatment. Front Endocrinol (Lausanne) 2022;13(1):1020827. [Google Scholar] | [Crossref]
- Bartkowiak-Wieczorek J, Jaros A, Gajdzińska A, Wojtyła-Buciora P, Szymański I, Szymaniak J, Janusz W, Walczak I, Jonaszka G, Bienert A. The Dual Faces of Oestrogen:The Impact of Exogenous Oestrogen on the Physiological and Pathophysiological Functions of Tissues and Organs. Int J Mol Sci 2024;25(15):8167. [Google Scholar] | [Crossref]
- Da Silva ID, Wuidar V, Zielonka M, Pequeux C. Unraveling the Dynamics of Estrogen and Progesterone Signaling in the Endometrium:An Overview. Cells 2024;13(15):1236. [Google Scholar] | [Crossref]
- Harding AT, Heaton NS. The Impact of Estrogens and Their Receptors on Immunity and Inflammation during Infection. Cancers (Basel) 2022;14(4):909. [Google Scholar] | [Crossref]
- Fedotcheva TA, Fedotcheva NI, Shimanovsky NL. Progesterone as an Anti-Inflammatory Drug and Immunomodulator:New Aspects in Hormonal Regulation of the Inflammation. Biomolecules 2022;12(9):1299. [Google Scholar] | [Crossref]
- Mikhalev V, Safonov V, Shabunin S, Parshin P, Sineva A, Lukina V. Ovarian hypofunction and its relationship to serum hormonal and cytokine profile in cattle. Adv Anim Vet Sci 2022;10(1):20-26. [Google Scholar] | [Crossref]
- Zhelavskyi M, Maryniuk M, Drobot M, Kostenko V, Boiko N, Paliukh T. Luteal insufficiency in canines:Assessment of progesterone dynamics and efficacy of combined hormonal treatment. Open Vet J 2025;15(6):2671-2681. [Google Scholar] | [Crossref]
- Mesen TB, Young SL. Progesterone and the luteal phase:a requisite to reproduction. Obstet Gynecol Clin North Am 2015;42(1):135-151. [Google Scholar] | [Crossref]
- Duncan WC. The inadequate corpus luteum. Reprod Fertil 2021;2(1):C1-C7. [Google Scholar] | [Crossref]
- Dong M, Dong Y, Bai J, Li H, Ma X, Li B, Wang C, Li H, Qi W, Wang Y, Fan A, Han C, Xue F. Interactions between microbiota and cervical epithelial, immune, and mucus barrier. Front Cell Infect Microbiol 2023;13(1):1124591. [Google Scholar] | [Crossref]
- Ismail N, Myint K, Khaing SL, Giribabu N, Salleh N. Cervical fluid pH, electrolytes and osmolarity changes and expression of ion transporters (ENaC, CFTR and AQP) in cervix of women with primary unexplained infertility. Mol Biol Rep 2023;50(8):6729-6737. [Google Scholar] | [Crossref]
- Dai T, Ma Z, Guo X, Wei S, Ding B, Ma Y, Dan X. Study on the Pattern of Postpartum Uterine Involution in Dairy Cows. Animals (Basel) 2023;13(23):3693. [Google Scholar] | [Crossref]
- Yasuda K, Yoshida A, Okada H. Conflicting Nongenomic Effects of Progesterone in the Myometrium of Pregnant Rats. Int J Mol Sci 2021;22(4):2154. [Google Scholar] | [Crossref]
- Sanchez L, Campos-Chillon F, Sargolzaei M, Peterson DG, Sprayberry KA, McArthur G, Anderson P, Golden B, Pokharel S, Abo-Ismail MK. Molecular Mechanisms Associated with the Development of the Metritis Complex in Dairy Cattle. Genes (Basel) 2024;15(4):439. [Google Scholar] | [Crossref]
- Sobstyl A, Chałupnik A, Mertowska P, Grywalska E. How Do Microorganisms Influence the Development of Endometriosis?Participation of Genital, Intestinal and Oral Microbiota in Metabolic Regulation and Immunopathogenesis of Endometriosis. Int J Mol Sci 2023;24(13):10920. [Google Scholar] | [Crossref]
- Podfigurna-Stopa A, Czyzyk A, Grymowicz M, Smolarczyk R, Katulski K, Czajkowski K, Meczekalski B. Premature ovarian insufficiency:the context of long-term effects. J Endocrinol Invest 2016;39(9):983-990. [Google Scholar] | [Crossref]
- Qin X, Du J, He R, Li Y, Li H, Liang X. Potential mechanisms and therapeutic strategies for LPS-associated female fertility decline. J Assist Reprod Genet 2024;41(10):2739-2758. [Google Scholar] | [Crossref]
- Kobayashi H, Nishio M, Umetani M, Shigetomi H, Imanaka S, Hashimoto H. Endometrial Aging and Reproductive Decline:The Central Role of Mitochondrial Dysfunction. Int J Mol Sci 2025;26(11):5060. [Google Scholar] | [Crossref]
- Dahiya S, Kumari S, Rani P, Onteru SK, Singh D. Postpartum uterine infection &ovarian dysfunction. Indian J Med Res 2018;148((Suppl)):S64-S70. [Google Scholar] | [Crossref]
- Kasimanickam R, Bhowmik P, Kastelic J, Ferreira J, Kasimanickam V. From Infection to Infertility:Diagnostic, Therapeutic, and Molecular Perspectives on Postpartum Metritis and Endometritis in Dairy Cows. Animals (Basel) 2025;15(19):2841. [Google Scholar] | [Crossref]
- Sheldon IM, Cronin JG, Bromfield JJ. Tolerance and Innate Immunity Shape the Development of Postpartum Uterine Disease and the Impact of Endometritis in Dairy Cattle. Annu Rev Anim Biosci 2019;7(1):361-384. [Google Scholar] | [Crossref]
- Głodkowska K, Brodzki P, Krakowski L, Wójcik A. Immune system of the uterus in cows and its functions –selected issues. Med Weter 2025;81(9):452-461. [Google Scholar] | [Crossref]
- Maheux-Lacroix S, Dodin S, Moore L, Bujold E, Lefebvre J, Bergeron MÈ. Preovulatory uterine flushing with saline as a treatment for unexplained infertility:a randomised controlled trial protocol. BMJ Open 2016;6(1):e009897. [Google Scholar] | [Crossref]
- Zangeneh FZ, Hantoushzadeh S. The physiological basis with uterine myometrium contractions from electro-mechanical/hormonal myofibril function to the term and preterm labor. Heliyon 2023;9(11):e22259. [Google Scholar] | [Crossref]
- Nybo L, Rasmussen P, Sawka MN. Performance in the heat-physiological factors of importance for hyperthermia-induced fatigue. Compr Physiol 2014;4(2):657-689. [Google Scholar] | [Crossref]
- Tindal K, Filby CE, Cousins FL, Ellery SJ, Vollenhoven B, Palmer K, Gordon A, Gargett CE, Davies-Tuck M. The composition of menstrual fluid, its applications, and recent advances to understand the endometrial environment:A narrative review. F &S Rev 2024;5(3):100075. [Google Scholar] | [Crossref]
- Katila T. Post-mating inflammatory responses of the uterus. Reprod Domest Anim 2012;47((Suppl 5)):31-41. [Google Scholar] | [Crossref]
- Satué K, La Fauci D, Medica P, Gímenez ED, Cravana C, Fazio E. Shifts between proinflammatory and anti-inflammatory profiles in pregnant mares:a review of physiological functions. Front Vet Sci 2025;12(1):1660759. [Google Scholar] | [Crossref]
- Li Q, Liu J, Nashun B, Bai S, Guo X, Wu L, Bao T. Inhibition of LPS-induced inflammation and signaling pathways in fallopian tube epithelial cells by estrogen and progesterone. BMC Vet Res 2025;21(1):421. [Google Scholar] | [Crossref]
- Magata F. Lipopolysaccharide-induced mechanisms of ovarian dysfunction in cows with uterine inflammatory diseases. J Reprod Dev 2020;66(4):311-317. [Google Scholar] | [Crossref]
- Choudhary S, Khan S, Rustagi S, Rajpal VR, Khan NS, Kumar N, Thomas G, Pandey A, Hamurcu M, Gezgin S, Zargar SM, Khan MK. Immunomodulatory Effect of Phytoactive Compounds on Human Health:A Narrative Review Integrated with Bioinformatics Approach. Curr Top Med Chem 2024;24(12):1075-1100. [Google Scholar] | [Crossref]
- Somagond YM, Das P, Dang AK, Yadav DK, Kittur PM, Panda BSK, Devi P, Kamboj A, Alhussien MN. Parenteral micronutrient supplementation enhances mammary immune function and colostrum-milk quality by modulating cytokine profiles and oxidative stress in transition crossbred cows. Front Immunol 2025;16(1):1669246. [Google Scholar] | [Crossref]
- Wu R, Li X, Li Z, Luo J, Zhang Z, Guo M. Selenium and Mammalian Uterine Health:A Comprehensive Review. Biology (Basel) 2025;14(9):1295. [Google Scholar] | [Crossref]
- Gawish AS, ElMofty MS, Jambi S, Felemban D, Ragheb YS, Elsayed SA. Phytotherapy in periodontics as an effective and sustainable supplemental treatment:a narrative review. J Periodontal Implant Sci 2024;54(4):209-223. [Google Scholar] | [Crossref]
- Khan MZ, Chen W, Liu X, Kou X, Khan A, Khan RU, Zahoor M, Wang C. An Overview of Bioactive Compounds'Role in Modulating the Nrf2/Keap1/NF-κB Pathway to Alleviate Lipopolysaccharide-Induced Endometritis. Int J Mol Sci 2024;25(19):10319. [Google Scholar] | [Crossref]
- Di Renzo L, De Lorenzo A, Fontanari M, Gualtieri P, Monsignore D, Schifano G, Alfano V, Marchetti M. Immunonutrients involved in the regulation of the inflammatory and oxidative processes:implication for gamete competence. J Assist Reprod Genet 2022;39(4):817-846. [Google Scholar] | [Crossref]
- Susilo J, Setyawan EMN, Hartanto S, Wibowo MH, Budiyanto A. Effect of GnRH treatment as a potential solution for ovarian disorders in dairy cows infected with foot and mouth disease in Indonesian smallholder farms. Open Vet J 2024;14(8):2079-2084. [Google Scholar] | [Crossref]
- Martire FG, Costantini E, D'Abate C, Schettini G, Sorrenti G, Centini G, Zupi E, Lazzeri L. Endometriosis and Adenomyosis:From Pathogenesis to Follow-Up. Curr Issues Mol Biol 2025;47(5):298. [Google Scholar] | [Crossref]
- Muhammad YA. Reproductive aging in biological females:mechanisms and immediate consequences. Front Endocrinol (Lausanne) 2025;16(1):1658592. [Google Scholar] | [Crossref]
- Diaz-Lundahl S, Garmo RT, Gillund P, Klem TB, Waldmann A, Krogenæs AK. Prevalence, risk factors, and effects on fertility of cytological endometritis at the time of insemination in Norwegian Red cows. J Dairy Sci 2021;104(6):6961-6974. [Google Scholar] | [Crossref]
- Yan X, Jiao J, Wang X. The pathogenesis, diagnosis, and treatment of chronic endometritis:a comprehensive review. Front Endocrinol (Lausanne) 2025;16(1):1603570. [Google Scholar] | [Crossref]
- Wang R, Watson A, Johnson N, Cheung K, Fitzgerald C, Mol BWJ, Mohiyiddeen L. Tubal flushing for subfertility. Cochrane Database Syst Rev 2020;10(10):CD003718. [Google Scholar] | [Crossref]
- Barabás K, Szabó-Meleg E, Ábrahám IM. Effect of Inflammation on Female Gonadotropin-Releasing Hormone (GnRH) Neurons:Mechanisms and Consequences. Int J Mol Sci 2020;21(2):529. [Google Scholar] | [Crossref]
- Jain V, Chodankar RR, Maybin JA, Critchley HOD. Uterine bleeding:how understanding endometrial physiology underpins menstrual health. Nat Rev Endocrinol 2022;18(5):290-308. [Google Scholar] | [Crossref]
- Lokshin V, Askar Y, Rybina A, Abshekenova A, Karibayeva S, Valiev R, Saduakas A. Antibiotics and Uterine Flushing versus Antibiotics Alone for Chronic Endometritis with Thin Endometrium in Assisted Reproductive Technology:A Single-Center Retrospective Cohort Study. Int J Fertil Steril 2025;19(2):186-192. [Google Scholar] | [Crossref]
- Zachut M, Tam J, Contreras GA. Modulating immunometabolism in transition dairy cows:the role of inflammatory lipid mediators. Anim Front 2022;12(5):37-45. [Google Scholar] | [Crossref]
- Luo X, Guo Y, Xu L. Microbial harmony in female reproductive health:exploring the impact of intestinal flora on ovarian function and disease pathogenesis. Am J Transl Res 2024;16(11):6274-6288. [Google Scholar] | [Crossref]
- Abdelli A, Raboisson D, Kaidi R, Ibrahim B, Kalem A, Iguer-Ouada M. Elevated non-esterified fatty acid and β-hydroxybutyrate in transition dairy cows and their association with reproductive performance and disorders:A meta-analysis. Theriogenology 2017;93(1):99-104. [Google Scholar] | [Crossref]
- Leziak A, Lipina J, Reclik M, Kocelak P. Dietary Modulation of Metabolic Health:From Bioactive Compounds to Personalized Nutrition. Metabolites 2025;15(9):624. [Google Scholar] | [Crossref]
- Mousaie A, Ghaedifard MR, Azizpour R, Sadeghi AA. Energy Intake During the Transition Period Triggers Substantial Impacts on Body Weight, Milk Production Efficiency, Milk Composition and Blood Biochemical Constituents in Fat-Tailed Ewes. Vet Med Sci 2025;11(6):e70644. [Google Scholar] | [Crossref]
- Llewellyn S, Fitzpatrick R, Kenny DA, Murphy JJ, Scaramuzzi RJ, Wathes DC. Effect of negative energy balance on the insulin-like growth factor system in pre-recruitment ovarian follicles of post partum dairy cows. Reproduction 2007;133(3):627-639. [Google Scholar] | [Crossref]
- Hu H, Zhang J, Xin X, Jin Y, Zhu Y, Zhang H, Fan R, Ye Y, Li D. Efficacy of natural products on premature ovarian failure:a systematic review and meta-analysis of preclinical studies. J Ovarian Res 2024;17(1):46. [Google Scholar] | [Crossref]
- Calder PC. Omega-3 polyunsaturated fatty acids and inflammatory processes:nutrition or pharmacology?. Br J Clin Pharmacol 2013;75(3):645-662. [Google Scholar] | [Crossref]
- Pokorska-Niewiada K, Ziętek M, Szydłowska I, Ryterska K, Szczuko M. The Role of Antioxidant Minerals in the Pathophysiology and Treatment of Endometriosis-Systematic Review. Antioxidants (Basel) 2025;14(10):1238. [Google Scholar] | [Crossref]
- Yang X, He Z, Hu R, Yan J, Zhang Q, Li B, Yuan X, Zhang H, He J, Wu S. Dietary β-Carotene on Postpartum Uterine Recovery in Mice:Crosstalk Between Gut Microbiota and Inflammation. Front Immunol 2021;12(1):744425. [Google Scholar] | [Crossref]
- Lewis ED, Meydani SN, Wu D. Regulatory role of vitamin E in the immune system and inflammation. IUBMB Life 2019;71(4):487-494. [Google Scholar] | [Crossref]
- Zhao L, Li X, Jing Y, Tang L, Lin F, Zhang Y, Tang Y, Chen C, Yang J, Liu X, Chen J. Biomarkers and immunotherapy in endometrial cancer:mechanisms and clinical applications. Front Immunol 2025;16(1):1684549. [Google Scholar] | [Crossref]
- Gremese E, Bruno D, Varriano V, Perniola S, Petricca L, Ferraccioli G. Serum Albumin Levels:A Biomarker to Be Repurposed in Different Disease Settings in Clinical Practice. J Clin Med 2023;12(18):6017. [Google Scholar] | [Crossref]
- Kaya S, Merhan O, Kaçar C, Colak A, Bozukluhan K. Determination of ceruloplasmin, some other acute-phase proteins, and biochemical parameters in cows with endometritis. Vet World 2016;9(10):1056-1062. [Google Scholar] | [Crossref]
- Abramiuk M, Grywalska E, Małkowska P, Sierawska O, Hrynkiewicz R, Niedźwiedzka-Rystwej P. The Role of the Immune System in the Development of Endometriosis. Cells 2022;11(13):2028. [Google Scholar] | [Crossref]
- Abu-Raya B, Michalski C, Sadarangani M, Lavoie PM. Maternal Immunological Adaptation During Normal Pregnancy. Front Immunol 2020;11(1):575197. [Google Scholar] | [Crossref]
- Constantin NT, Bercea-Strugariu CM, Bîrţoiu D, Posastiuc FP, Iordache F, Bilteanu L, Serban AI. Predicting Pregnancy Outcome in Dairy Cows:The Role of IGF-1 and Progesterone. Animals (Basel) 2023;13(10):1579. [Google Scholar] | [Crossref]
- Horst EA, Kvidera SK, Baumgard LH. Invited review:The influence of immune activation on transition cow health and performance-A critical evaluation of traditional dogmas. J Dairy Sci 2021;104(8):8380-8410. [Google Scholar] | [Crossref]
- Martire FG, Costantini E, Ianes I, d'Abate C, De Bonis M, Capria G, Piccione E, Andreoli A. Nutrition and Uterine Fibroids:Clinical Impact and Emerging Therapeutic Perspectives. J Clin Med 2025;14(20):7140. [Google Scholar] | [Crossref]
- Song Y, Cheng J, Yu H, Wang Z, Bai Y, Xia C, Xu C. Early Warning for Ovarian Diseases Based on Plasma Non-esterified Fatty Acid and Calcium Concentrations in Dairy Cows. Front Vet Sci 2021;8(1):792498. [Google Scholar] | [Crossref]
- Mota LFM, Giannuzzi D, Pegolo S, Trevisi E, Ajmone-Marsan P, Cecchinato A. Integrating on-farm and genomic information improves the predictive ability of milk infrared prediction of blood indicators of metabolic disorders in dairy cows. Genet Sel Evol 2023;55(1):23. [Google Scholar] | [Crossref]
- Kia S, Mohri M, Seifi HA. Association of precalving serum NEFA concentrations with postpartum diseases and reproductive performance in multiparous Holstein cows:Cut-off values. Vet Med Sci 2023;9(4):1757-1763. [Google Scholar] | [Crossref]
- Suthar VS, Canelas-Raposo J, Deniz A, Heuwieser W. Prevalence of subclinical ketosis and relationships with postpartum diseases in European dairy cows. J Dairy Sci 2013;96(5):2925-2938. [Google Scholar] | [Crossref]
- Menzel A, Samouda H, Dohet F, Loap S, Ellulu MS, Bohn T. Common and Novel Markers for Measuring Inflammation and Oxidative Stress Ex Vivo in Research and Clinical Practice-Which to Use Regarding Disease Outcomes?. Antioxidants (Basel) 2021;10(3):414. [Google Scholar] | [Crossref]
- Pohl A, Burfeind O, Heuwieser W. The associations between postpartum serum haptoglobin concentration and metabolic status, calving difficulties, retained fetal membranes, and metritis. J Dairy Sci 2015;98(7):4544-4551. [Google Scholar] | [Crossref]
- Vilotić A, Nacka-Aleksić M, Pirković A, Bojić-Trbojević Ž, Dekanski D, Krivokuća MJ. IL-6 and IL-8:An Overview of Their Roles in Healthy and Pathological Pregnancies. Int J Mol Sci 2022;23(23):14574. [Google Scholar] | [Crossref]
- Valera H, Chen A, Grive KJ. The Hypothalamic–Pituitary–Ovarian Axis, Ovarian Disorders, and Brain Aging. Endocrinology 2025;166(10):bqaf137. [Google Scholar] | [Crossref]
- Walsh SW, Williams EJ, Evans AC. A review of the causes of poor fertility in high milk producing dairy cows. Anim Reprod Sci 2011;123((3-4)):127-138. [Google Scholar] | [Crossref]
- Çolakoğlu HE, Yazlık MO, Pekcan M, Kaya U, Kaçar C, Vural MR, Kurt S, Yildirim MM, Bas A, Küplülü Ş. Impact of Prepartum Body Condition Score Loss on Metabolic Status During the Transition Period and Subsequent Fertility in Brown Swiss Dairy Cows. J Vet Res 2019;63(3):375-382. [Google Scholar] | [Crossref]
- LeBlanc S. Monitoring metabolic health of dairy cattle in the transition period. J Reprod Dev 2010;56((Suppl)):S29-S35. [Google Scholar] | [Crossref]
- Moustakli E, Stavros S, Katopodis P, Potiris A, Drakakis P, Dafopoulos S, Zachariou A, Dafopoulos K, Zikopoulos K, Zikopoulos A. Gut Microbiome Dysbiosis and Its Impact on Reproductive Health:Mechanisms and Clinical Applications. Metabolites 2025;15(6):390. [Google Scholar] | [Crossref]
- Frountzas M, Stefanoudakis D, Mela E, Theodorou P, Theodoropoulos GE, Theodorou D, Toutouzas KG. Peripheral Parenteral Nutrition and Personalized Nutritional Approach After Colorectal Resection Surgery:A Comprehensive Review of Current Evidence. Medicina (Kaunas) 2025;61(8):1459. [Google Scholar] | [Crossref]
- van Loenen MR, Remie LB, van Trijp MPH, Jansen MG, Marques JP, Claassen JAHR, van de Rest O, Vermeiren Y, Smidt N, Sikkes SAM, Deckers K, Zwan MD, van der Flier WM, Köhler S, Steegenga WT, Oosterman JM, Aarts E. The Effects of a Multidomain Lifestyle Intervention on Brain Function and Its Relation With Immunometabolic Markers and Intestinal Health in Older Adults at Risk of Cognitive Decline:Study Design and Baseline Characteristics of the HELI Randomized Controlled Trial. JMIR Res Protoc 2025;14(1):e69814. [Google Scholar] | [Crossref]
- Arshad R, Gulshad L, Haq IU, Farooq MA, Al-Farga A, Siddique R, Manzoor MF, Karrar E. Nanotechnology:A novel tool to enhance the bioavailability of micronutrients. Food Sci Nutr 2021;9(6):3354-3361. [Google Scholar] | [Crossref]
- El Safadi D, Paulo-Ramos A, Hoareau M, Roche M, Krejbich-Trotot P, Viranaicken W, Lebeau G. The Influence of Metabolism on Immune Response:A Journey to Understand Immunometabolism in the Context of Viral Infection. Viruses 2023;15(12):2399. [Google Scholar] | [Crossref]
- Altanam SY, Darwish N, Bakillah A. Exploring the Interplay of Antioxidants, Inflammation, and Oxidative Stress:Mechanisms, Therapeutic Potential, and Clinical Implications. Diseases 2025;13((9)):309. [Google Scholar] | [Crossref]
- Miah S, Chy WR, Ahmed T, Suchi M, Muhtady A, Ahmad SNU, Hossain MA. Emerging trends in nanotechnologies for vitamin delivery:Innovation and future prospects. Nano Trends 2025;10(1):100126. [Google Scholar] | [Crossref]
- Boni R, Gualandi SC. Relationship between Oxidative Stress and Endometritis:Exploiting Knowledge Gained in Mares and Cows. Animals (Basel) 2022;12(18):2403. [Google Scholar] | [Crossref]
- Parker J, O'Brien C, Uppal T, Tremellen K. Molecular Impact of Metabolic and Endocrine Disturbance on Endometrial Function in Polycystic Ovary Syndrome. Int J Mol Sci 2025;26(20):9926. [Google Scholar] | [Crossref]
- Limam I, Abdelkarim M, Kacem-Berjeb K, Khrouf M, Feki A, Braham M, Chakroun N. Precision Therapeutic and Preventive Molecular Strategies for Endometriosis-Associated Infertility. Int J Mol Sci 2025;26(16):7706. [Google Scholar] | [Crossref]