


ABSTRACT
Background and Aim: In
Materials and Methods: A total of 1,485 high-quality buffalo oocytes were subjected to
Results: The nuclear maturation rate (metaphase II stage) was significantly higher (p < 0.01) in the IGF-1 and lycopene groups (85.2% and 87.3%, respectively) compared with the control (73.3%) and α-tocopherol groups (76.2%). Cleavage, morula, and blastocyst formation rates were also significantly higher (p < 0.01) in the IGF-1 (89.3%, 28.5%, and 20.6%) and lycopene (84.2%, 30.8%, and 32.7%) groups than in the control (75.1%, 20.3%, and 12.2%) and α-tocopherol (76.7%, 23.2%, and 14.4%) groups. Lycopene produced the highest blastocyst yield. Mitochondrial fluorescence intensity was significantly greater (p < 0.01) in all supplemented groups compared with the control. Diffuse mitochondrial distribution predominated in IGF-1- and lycopene-treated oocytes, indicating improved cytoplasmic competence and metabolic activity, whereas α-tocopherol treatment was associated with increased peripheral mitochondrial localization.
Conclusion: Supplementation of IVM medium with 100 ng/mL IGF-1 or 0.2 μM lycopene significantly enhances nuclear maturation, mitochondrial activity, and embryo developmental competence of buffalo oocytes. Lycopene demonstrated the most pronounced improvement in blastocyst formation, suggesting superior antioxidant protection during maturation. These findings highlight the importance of optimizing mitochondrial function and oxidative balance in buffalo IVEP systems and provide a potential strategy to improve reproductive biotechnology outcomes in buffalo.
Keywords: buffalo oocytes, embryo development, insulin-like growth factor-1,
INTRODUCTION
Buffaloes (
Among these technologies,
To mitigate oxidative stress and improve oocyte quality, various supplements have been incorporated into IVM media. These supplements may include hormones, growth factors, and antioxidants that modulate cellular metabolism and protect oocytes from oxidative damage. Growth factors such as insulin-like growth factor-1 (IGF-1) and antioxidants such as lycopene and α-tocopherol have attracted considerable attention because of their ability to enhance oocyte maturation, fertilization success, and embryo developmental competence. Nevertheless, most previous studies have evaluated these supplements individually. Investigations involving IGF-1 have primarily been conducted in buffalo and bovine species, whereas lycopene supplementation has mainly been studied in cattle and pigs. Similarly, α-tocopherol has been examined primarily in cattle, sheep, and porcine models. Consequently, comparative evaluations of these supplements under identical IVC conditions in buffalo oocytes remain scarce.
The addition of growth factors and antioxidants to embryo culture media has been shown to enhance blastocyst development, increase hatching rates, and improve pregnancy outcomes in several livestock species [4]. Among these supplements, IGF-1 is considered one of the most important growth factors involved in ovarian follicular development and early embryogenesis. IGF-1 acts as a potent anti-apoptotic factor during preimplantation embryo development in buffalo (
Lycopene is another promising supplement for improving oocyte competence. It is a naturally occurring carotenoid characterized by a highly conjugated molecular structure consisting of 13 double bonds, 11 of which are conjugated. This structure confers exceptionally strong antioxidant properties, making lycopene approximately twice as effective as β-carotene in scavenging free radicals [9, 10]. Lycopene has been shown to enhance mitochondrial activity, reduce apoptosis, and increase both the inner cell mass (ICM) and total cell number in bovine blastocysts [11–13]. Residiwati
Another important antioxidant used in reproductive biotechnology is α-tocopherol, the biologically active form of vitamin E. As a lipid-soluble antioxidant, α-tocopherol protects cellular membranes from oxidative damage by scavenging free radicals and preventing lipid peroxidation [15]. A previous study has reported that supplementation of buffalo oocytes with 100 μM vitamin E during IVM increases maturation, cleavage, morula, and blastocyst rates [16]. In bovine embryos, α-tocopherol has also been shown to improve embryonic quality during IVM [17]. Furthermore, α-tocopherol supplementation enhances granulosa cell viability in pigs [18], supports folliculogenesis and oocyte competence in cattle [15], and improves embryonic development in gilts [19] and sheep [20]. Vitamin E also prevents membrane damage [21], inhibits lipid peroxidation in bovine cells [22], and reduces apoptosis in mammalian oocytes [23].
Mitochondria play a fundamental role in regulating oocyte maturation and embryonic development. These organelles serve as the primary source of cellular energy in the form of adenosine triphosphate (ATP), which is essential for transcriptional and translational processes occurring during oocyte maturation [24]. During maturation, mitochondrial DNA copy number increases progressively, and mitochondrial distribution patterns within the oocyte cytoplasm undergo substantial changes [24]. The proper distribution and activity of mitochondria are therefore essential for maintaining oocyte developmental competence.
Growth factors and antioxidants are known to modulate mitochondrial function in
These protective mechanisms ultimately lead to improved IVM rates and enhanced embryo developmental competence in several livestock species [28]. Mature metaphase II (MII) oocytes typically contain approximately 500,000 copies of mitochondrial DNA, which are essential for maintaining mitochondrial activity and regulating intracellular oxidative balance [29]. The spatial distribution of mitochondria within the oocyte cytoplasm is tightly regulated and plays a crucial role in cellular organization and metabolic activity [30].
Recent studies have highlighted the importance of mitochondria-targeted antioxidants, which can accumulate within mitochondria at concentrations 100–1,000 times higher than those found in the surrounding cytoplasm [31]. Monitoring mitochondrial distribution patterns has therefore emerged as a valuable biological indicator for assessing oocyte quality and developmental competence [32]. During meiosis I (MI), mitochondria gradually surround the developing spindle apparatus and form clusters within the cytoplasm. By the MII stage, mitochondria become widely dispersed throughout the cytoplasm, with higher concentrations observed in the spindle hemisphere and cortical regions of the oocyte [33]. Nevertheless, considerable variability exists in the intensity and distribution of mitochondria among oocytes of different species and developmental stages.
Despite increasing recognition of the importance of mitochondrial dynamics in oocyte maturation, a comprehensive characterization of mitochondrial distribution patterns during buffalo oocyte maturation remains limited. Moreover, objective and quantitative approaches to evaluating mitochondrial distribution in oocytes remain underdeveloped [32].
Despite substantial advances in reproductive biotechnology, the efficiency of buffalo
Therefore, the present study aimed to compare the effects of IGF-1, lycopene, and α-tocopherol supplementation during IVM on the developmental competence of buffalo (
MATERIALS AND METHODS
Ethical approval
All procedures involving animal biological materials were performed in strict accordance with national and international guidelines for the ethical use of animals in research. This study did not involve any live experimental animals; ovaries were sourced exclusively from routinely slaughtered adult buffaloes (
Study period and location
The study was conducted during the buffalo breeding season, from October to March, between 2022 and 2025. The
Collection and transport of the buffalo ovaries
Ovaries were collected from clinically healthy, non-pregnant adult buffaloes (
Retrieval and grading of buffalo oocytes
Follicular aspiration
Cumulus–oocyte complexes (COCs, n = 4,250 oocytes) were aspirated from 2–8 mm follicles, and only excellent and good quality oocytes were selected, whereas fair and denuded (n = 1,342 oocytes) were excluded. COCs were retrieved from follicles that ranged in diameter from 2 to 8 mm using a sterile disposable 20 mL syringe with an 18-gauge needle containing 1 mL of phosphate-buffered saline (PBS) supplemented with 6 mg/mL of bovine serum albumin (fraction V) and 50 μg/mL of gentamicin. After aspiration, the follicular fluid was placed in a 15-mL sterile Falcon tube and allowed to settle in a water bath at 37°C for 15 min.
Washing and handling
The COCs were washed three times in the aspiration medium to remove blood and debris. Handling was performed carefully to preserve oocyte integrity.
Grading criteria
The quality of the COCs was examined using a stereomicroscope (Zeiss, Oberkochen, Germany) at 90× magnification [35]. The quality of the oocytes was classified into four categories based on the condition of the cumulus layers and the homogeneity of the cytoplasm: excellent, good, fair, and denuded [36]. Excellent: Oocytes with more than five layers of cumulus cells and dark cytoplasm with uniform granulation. Good: Dark cytoplasm with uniform granulation and three to five layers of cumulus cells. Fair: Oocytes that are partially enclosed by cumulus cells, with minimally granulated cytoplasm. Denuded: Oocytes that lack cumulus cells and are only covered by zona pellucida [36, 37]. Only excellent and good quality oocytes (n = 2908) were selected for subsequent experiments, whereas fair and denuded oocytes (n = 1342) were excluded. Excellent oocytes = 1650 (38.82%), good oocytes = 1,258 (29.6%), fair oocytes = 770 (18.12%), and denuded oocytes = 572 (13.46%).
IVM of buffalo oocytes
Composition of the maturation medium
The basic maturation medium consisted of TCM-199 (M4530, Sigma-Aldrich, St. Louis, MO, USA) with 10% fetal calf serum (FCS) + 10 μg/mL follicle-stimulating hormone (FSH) (Follitropin-V, Bioniche Animal Health, Belleville, ON, Canada) + 50 μg/mL gentamicin and filtered using 0.2 μm syringe filter (Thermo Fisher Scientific, Waltham, MA, USA) and incubated for at least 2 h in a humidified atmosphere (95%) under 5% CO2 at 38.5°C before culturing of the oocytes. Excellent and good oocytes
Preparation of IGF-1, lycopene, and α-tocopherol
The dose and preparation of IGF-1 are according to Ismail The dose and preparation of lycopene were performed according to Residiwati The dose and preparation of α-tocopherol were as described by Thiyagarajan and Valivittan [16]. Preparation of vit E as a molar mass (430.7) g/L and vit E as a fat-soluble stock solution, each 1 mL contain 0.95 g; addition of 1 μL of vit E to 219 μL of basic maturation media for a concentration of 10000 μM, then final working solution is 10 μL from 220 and added to 990 mL maturation media for a concentration of 100 μM.
Culture conditions
Assessment of cytoplasmic maturation
Cytoplasmic maturation was assessed in all
Assessment of nuclear maturation
Nuclear maturation evaluation was based on the presence of the first polar body (1st PB) in the oocyte, which was determined after gently removing the cumulus cells of all
In vitro fertilization (IVF) of buffalo oocytes
Semen thawing and washing
Frozen buffalo semen was purchased from the Abbasia Artificial Insemination Center, General Organization for Veterinary Services, Ministry of Agriculture, Egypt. The same batch of the same bull with proven fertility was used throughout the study (Egy. Buff. ASWAD 656 ABASSIA AIC). A 0.25 mL straw of frozen semen was thawed in a water bath at 37°C for 30 s. The spermatozoa (60%–70% progressive motility) were washed with 3 mL of sperm Tyrode’s albumin lactate pyruvate medium supplemented with 1 μg/mL heparin, 3 mg/mL BSA, 2.5 mg/mL hypotaurine and 50 μg/mL gentamicin. The mixture was then centrifuged at 1,800 rpm (approximately 700 ×
Sperm concentration adjustment
Following these two centrifugation steps, the remaining semen pellet was combined with 200 μL Fert-TALP medium, and the sperm number was counted using a hemocytometer. The final sperm concentration was modified to 1 × 106 spermatozoa/mL before being placed in the four-well plate.
Insemination procedure
Sperm suspension with 300 μL Fert-TALP medium was placed into a four-well culture plate and covered with 200 μL of warm mineral oil. Both oocytes and sperm were cultured in a CO2 incubator with a 5% CO2 humidified atmosphere at 38.5°C for 18 h.
Fertilization assessment
Fertilization was assessed based on the presence of the second PB. The fertilization rate was evaluated according to established guidelines [3]. Polyspermy-fertilized oocytes were excluded from assessment and IVC for development.
IVC of buffalo oocytes
Culture medium composition
Modified synthetic oviduct fluid medium (mSOF) supplemented with 5 mg/mL BSA and 50 μg/mL gentamicin. Fertilized oocytes were cultured in a four-well culture plate using mSOF medium (50 oocytes/500 μL medium) for 7 days (change medium every 2 days) in a humidified atmosphere (95%) under 5% CO2 at 38.5°C. A fresh culture medium was introduced every two days for a total of seven days.
Embryo staging and evaluation
Cleavage and embryo development rates were assessed on days 2, 5, and 7 using an inverted microscope (Zeiss). The embryo cleavage, morula, and early blastocyst rates were evaluated according to
Mitochondrial staining and confocal microscopy: Fixation and permeabilization
The
The staining procedure
Mito Tracker Red FM stain (Thermo Fisher Scientific), a fluorescent marker specific to mitochondria, was used to stain mature buffalo oocytes
Imaging parameters
A confocal microscope with four laser beams (Zeiss LSM 710) and Zen 2010 software (https://www.zeiss. com/microscopy/us/products/software/zeiss-zen-lite.html) were used to image the stained oocytes. A confocal microscope connected to a P-TMP unit was used to take images without fluorescence; then the red and blue laser beams were used to image red mitochondria and blue for nuclear DNA and the 1st PB. The confocal microscope was calibrated using the standard stained slide that came with the microscope to ensure the laser beams were functioning properly before imaging. We selected the black oocytes and white under 200× magnification. Then, the laser beams were used to capture images of the oocytes. The appearance of the nucleus and the first polar body (PB), which should be stained with DAPI to show blue fluorescence, along with the red image of the mitochondria, are the key indicators for imaging. The DAPI stain was examined using excitation and emission wavelengths of 358 and 460 nm, respectively. The Mito Tracker Red stain was analyzed using an emission filter set to 644 nm and an excitation wavelength of 581 nm. An optical section was captured at the plane where the nucleus was visible for each oocyte.
Quantification and distribution criteria
A confocal microscope (Zeiss LSM 710) was used to assess the mitochondrial distribution and morphology of the stained oocytes. The distribution of mitochondria in oocytes was categorized as follows: (1) peripheral distribution, where mitochondria were found on the inner surface of the zona pellucida; (2) semi-peripheral distribution, where mitochondria were found in the inner region of the oocyte but were distributed unevenly; (3) diffuse distribution, characterized by a homogeneous arrangement of mitochondria throughout the entire oocyte; and (4) semi-diffused distribution, where mitochondria were concentrated in the center with an uneven distribution toward the oocyte wall. The confocal microscope’s software automatically detected mitochondrial intensity [38]. Each group of
Experimental design and replication
The excellent and good oocytes were randomly allocated during IVM and culture, 50 oocytes in 500 μL media/well of a four-well tissue culture plate. The oocytes were randomly distributed into four experimental groups: control (no supplementation), IGF-1 (100 ng/mL), lycopene (0.2 μM), and α-tocopherol (100 μM). Each experiment was replicated ten times using independent oocyte batches. Oocytes were randomly allocated into the control, IGF-1, lycopene, and α-tocopherol groups. Each experiment was replicated 10 times. The sample sizes were based on power analysis to detect significant differences (power = 0.8, α = 0.05). Experiments were conducted on separate days, and Figure 1 summarizes the number of replicates and oocytes analyzed per group.
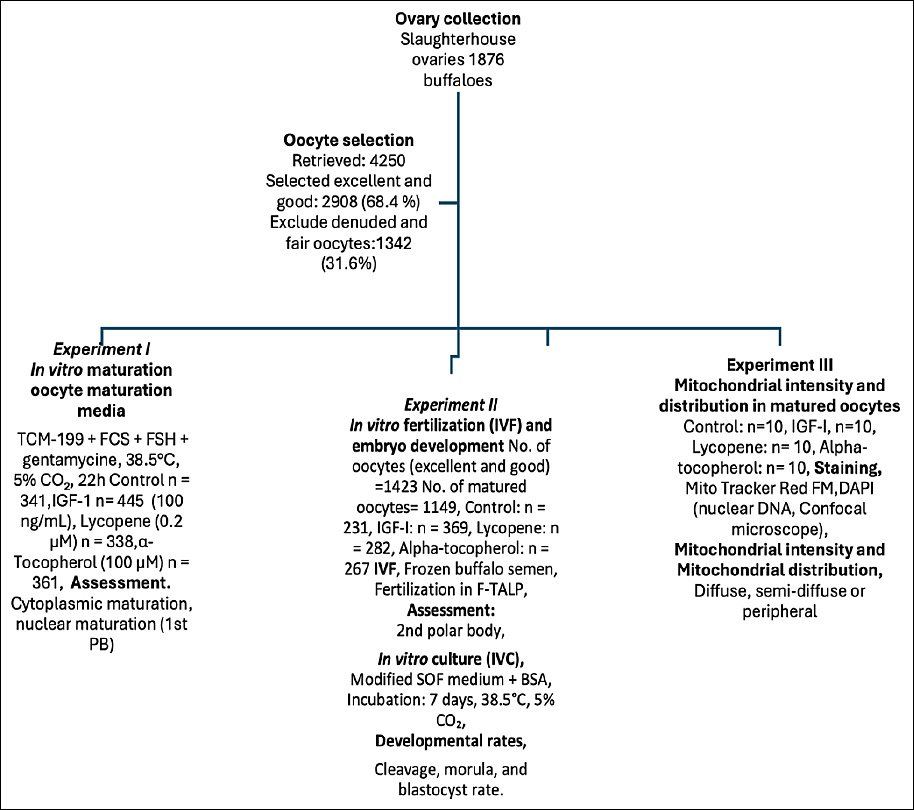
Figure 1. Schematic overview of the experimental workflow for
Experiment I: Effects of IGF-1, lycopene, and α-tocopherol on IVM of buffalo oocytes
Excellent and good oocytes (n = 1485) were matured
Experiment II: Effect of IGF-1, lycopene, and α-tocopherol on buffalo oocyte development
After the
Experiment III: Effect of IGF-1, lycopene, and α-tocopherol on intensity and distribution in in vitro-matured buffalo oocytes
The four groups of
Statistical analysis
Statistical analysis was performed using the SPSS version 28.0 software (SPSS Inc., Chicago, IL, USA). Data were tested for normality before analysis. Quantitative data are expressed as mean ± standard error. Differences among groups were evaluated using one-way analysis of variance followed by Tukey’s post hoc test, while proportional data were analyzed using the chi-square test or Fisher’s exact test [39]. Differences were considered statistically significant at p ≤ 0.05.
RESULTS
Experiment I: Effect of IGF-1, lycopene, and α-tocopherol on IVM rates of buffalo oocytes
Cumulus cell expansion rate
The effect of the addition of 100 ng/mL IGF-1 (IGF-1 group), 0.2 μM lycopene (lycopene group), and 100 μM α-tocopherol (α-tocopherol group) to IVM medium (TCM-199) on cumulus cell expansion of
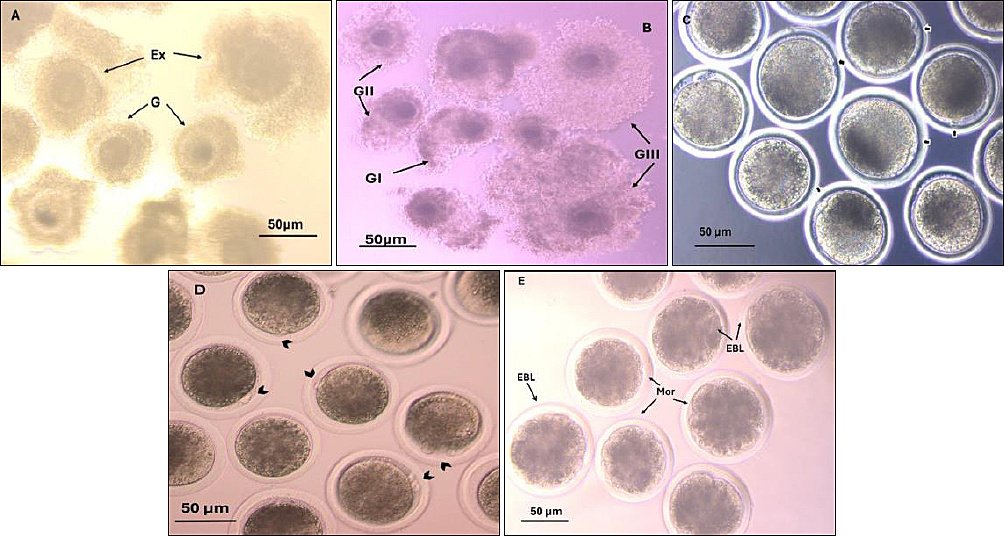
Figure 2. Buffalo oocyte quality and oocyte developmental competence. A) Oocyte quality (G=good, Ex = excellent). B) Cumulus expansion (GIII =full expansion, GII = moderate expansion, GI = little expansion). C) Nuclear matured oocytes, the arrow shows the 1st polar body. D) Fertilized oocytes, the arrow shows the 2nd polar body. F) Transferable embryo (Mor = morula, EBL = early blastocyst). These pictures were taken using a Zeiss inverted microscope at 200× magnification, with a scale of 50 μm.
Table 1. Effect of IGF-1, lycopene, and α-tocopherol supplementation on cumulus cell expansion (mean ± standard error and percentage) of
| Groups | Total oocytes | GIII (No.) | GIII (Mean ± SE) | GIII (%) | GII (No.) | GII (Mean ± SE) | GII (%) | GI (No.) | GI (Mean ± SE) | GI (%) | G0 (No.) | G0 (Mean ± SE) | G0 (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 341 | 141 | 41.34 ± 0.46b | 41.3b | 57 | 16.76 ± 0.40b | 16.7b | 57 | 16.81 ± 0.73b | 16.7b | 86 | 25.07 ± 0.52a | 25.2a |
| IGF-1 | 445 | 186 | 41.77 ± 0.32b | 41.8b | 75 | 16.82 ± 0.30b | 16.9b | 89 | 20.01 ± 0.32a | 20.0a | 95 | 21.42 ± 0.30b | 21.3b |
| Lycopene | 338 | 196 | 58.00 ± 0.46a | 58.0a | 97 | 28.63 ± 0.49a | 28.7a | 28 | 8.44 ± 0.62c | 8.3c | 17 | 5.04 ± 0.39c | 5.0c |
| α-tocopherol | 361 | 155 | 42.57 ± 0.82b | 42.9b | 64 | 17.62 ± 0.27b | 17.7b | 72 | 20.06 ± 0.37a | 19.9a | 70 | 19.75 ± 0.89b | 19.4b |
Replicates = 10 a,b,c Means within the same column bearing different superscripts differ significantly (p < 0.01). IGF-1 = insulin-like growth factor-1, ng = nanogram, μM = micromolar, SE = Standard error.
Nuclear maturation rate
Mature oocytes were examined for the presence of first Pb as an indication for meiosis II (Table 2, Figure 2C) and found that the maturation rate (mean ± SE) in the IGF-1 group and lycopene group (85.12 ± 0.46%, 87.07 ± 1.41%, respectively) was significantly (p < 0.01) higher than the TCM (73.34 ± 0.16%) or α-tocopherol group (76.27±0.78 %). The
Table 2. Effect of IGF-1, lycopene, and α-tocopherol supplementation on nuclear maturation rate (mean ± standard error and percentage) of
| Groups | Total oocytes | Maturation rate (1st PB) | Oocytes without a polar body rate | ||||
|---|---|---|---|---|---|---|---|
|
| |||||||
| No. | Mean ± SE (%) | % | No. | Mean ± SE (%) | % | ||
| Control | 341 | 250 | 73.34 ± 0.16b | 73.3b | 91 | 26.66 ± 0.16a | 26.7a |
| IGF-1 | 445 | 379 | 85.12 ± 0.46a | 85.2a | 66 | 14.89 ± 0.46b | 14.8b |
| Lycopene | 338 | 295 | 87.07 ± 1.41a | 87.3a | 43 | 12.93 ± 1.41b | 12.7b |
| α-Tocopherol | 361 | 275 | 76.27 ± 0.78b | 76.2b | 86 | 23.73 ± 0.78a | 23.8a |
Replicates = 10
* Total number of oocytes: number of excellent and good quality oocytes. a,b Means within the same column bearing different superscripts differ significantly (p < 0.01). IGF-1 = insulin-like growth factor-1, PB = polar body, ng = nanogram, μM = micromolar, SE = Standard error.
Experiment II: Effect of IGF-1, lycopene, and α-tocopherol on embryo development in buffalo
The results showed no significant difference (p > 0.05) between control, IGF-1, lycopene, and α-tocopherol groups (85.37 ± 0.61%, 88.03 ± 0.75%, 87.82±1.22%, and 87.99 ± 1.01%, respectively) in the fertilization rate (mean ± S. E). Cleavage rate, morula rate, and blastocyst rate (mean ± S. E) were significantly (p < 0.01) higher in the IGF-1 group (88.99 ± 0.57%, 28.52 ± 1.03% and 20.70 ± 0.42%, respectively) and lycopene group (84.33 ± 0.33%, 30.91 ± 0.84%, and 32.71 ± 0.36%, respectively) when compared with TCM group (75.11 ± 0.64%, 20.37 ± 0.48% and 12.31 ± 0.80%, respectively) and α-tocopherol group (76.51 ± 0.45%, 22.85 ± 0.73% and 14.28 ± 0.85%, respectively). The blastocyst rate (mean ± S. E) was significantly higher (p < 0.01) in the lycopene group than in the IGF-1 group (Table 3, Figure 2D and E). The results showed no significant difference (p > 0.05) in fertilization rate among the control, IGF-1, lycopene, and α-tocopherol groups (85.3%, 88.3%, 87.6%, and 88.4%, respectively). Cleavage rate and morula rate and blastocyst rate were significantly (p < 0.01) higher in the IGF-1 group (89.3%, 28.5%, and 20.6%, respectively) and lycopene group (84.2%, 30.8%, and 32.7%, respectively) than in the TCM group (75.10%, 20.3%, and 12.2%, respectively) and α-tocopherol group (76.7%,23.2% and 14.4% respectively). The blastocyst rate was significantly higher (p < 0.01) in the lycopene group than in the IGF-1 group (Table 3, Figures 2D and E).
Table 3. Effect of IGF-1, lycopene, and α-tocopherol supplementation on embryo development (mean ± standard error and percentage) in buffalo.
| Groups | No. of oocytes | Fertilization rate | Cleavage rate | Morula rate | Blastocyst rate | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| |||||||||||||
| No. | Mean ± SE (%) | % | No. | Mean ± SE (%) | % | No. | Mean ± SE (%) | % | No. | Mean ± SE (%) | % | ||
| Control | 231 | 197 | 85.37 ± 0.61a | 85.3a | 148 | 75.11 ± 0.64b | 75.1b | 30 | 20.37 ± 0.48b | 20.3b | 18 | 12.31 ± 0.80c | 12.2c |
| IGF-1 | 369 | 326 | 88.03 ± 0.75a | 88.3a | 291 | 88.99 ± 0.57a | 89.3a | 83 | 28.52 ± 1.03a | 28.5a | 60 | 20.70 ± 0.42b | 20.6b |
| Lycopene | 282 | 247 | 87.82 ± 1.22a | 87.6a | 208 | 84.33 ± 0.33a | 84.2a | 64 | 30.91 ± 0.84a | 30.8a | 68 | 32.71 ± 0.36a | 32.7a |
| α-tocopherol | 267 | 236 | 87.99 ± 1.01a | 88.4a | 181 | 76.51 ± 0.45b | 76.7b | 42 | 22.85 ± 0.73b | 23.2b | 26 | 14.28 ± 0.85c | 14.4c |
Replicates = 10
* Oocytes with first polar body. a,b,c Means within the same column bearing different superscripts differ significantly (p < 0.01). IGF-1 = insulin-like growth factor-1, PB = polar body, ng = nanogram, μM = micromolar.
Experiment III: Effect of IGF-1, lycopene, and α-tocopherol on mitochondrial intensity and distribution
Mitochondrial mean ± S. E intensity (Table 4) was significantly (p < 0.01) higher in the IGF-1 group (290.72 ± 10.19), lycopene group (248.33 ± 5.11), and the α-tocopherol group (208.03 ± 4.12) when compared with the TCM group (176 ± 2.71). The diffuse distribution of mitochondria, as shown in the IGF-1 and lycopene groups in Figure 3, was significantly (p < 0.01) higher in the IGF-1 and lycopene groups (90% and 70%, respectively) when compared with the α-tocopherol group (30%). The semi-diffused mitochondrial distribution (shown in the control group in Figure 3) of the α-tocopherol group was significantly (p < 0.01) higher (40%) than that of the IGF-1, lycopene, and control groups (30%, 30%, and 10%, respectively) (Table 5, Figure 3). Conversely, the peripheral distribution of mitochondria was observed in the α-tocopherol group in Figure 3 (30%), significantly (p < 0.01) higher than in the other groups (no peripheral distribution, 0.0%) (Table 5, Figure 3).
Table 4. Effect of IGF-1, lycopene, and α-tocopherol supplementation on mitochondrial intensity (mean ± standard error) of
| Groups | Number of oocytes imaged | Mean ± SE |
|---|---|---|
| Control | 30 | 176 ± 2.71d |
| IGF-1 | 30 | 290.72 ± 10.19a |
| Lycopene | 30 | 248.33 ± 5.11b |
| α-tocopherol | 30 | 208.03 ± 4.12c |
Replicates = 3. a,b,c,d Means within the same column bearing different superscripts differ significantly (
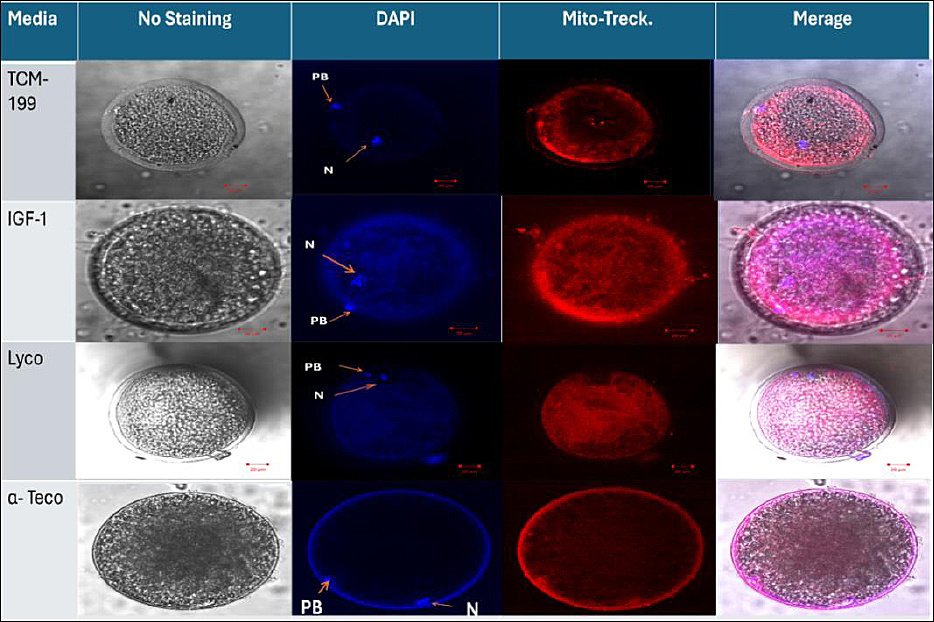
Figure 3. Detection of the effects of IGF-1, lycopene, and α-tocopherol on viability and mitochondrial distribution of
Table 5. Effect of IGF-1, lycopene, and α-tocopherol supplementation on mitochondrial distribution (number and percentage) of
| Groups | Matured oocytes | Diffused n (%) | Semi-diffused n (%) | Peripheral n (%) |
|---|---|---|---|---|
| Control | 30 | 27 (90)a*** | 3 (10)c** | 0 (0)b* |
| IGF-1 | 30 | 21 (70)b*** | 9 (30)b** | 0 (0)b* |
| Lycopene | 30 | 21 (70)b*** | 9 (30)b** | 0 (0)b* |
| α-tocopherol | 30 | 9 (30)c** | 12 (40)a*** | 9 (30)a* |
Replicates = 3. ***, **, * Significant difference within the same row (p < 0.01). a,b,c Means within the same column bearing different superscripts differ significantly (
DISCUSSION
Effect of IGF-1, lycopene, and α-tocopherol supplements to the maturation medium on the rate of maturation of buffalo oocytes
Cytoplasmic maturation
Our study revealed that lycopene improves the cytoplasmic maturation rate of buffalo oocytes when compared to the IGF-1, α-tocopherol, and control groups. Lycopene increased cytoplasmic maturation in buffalo oocytes due to its mechanism of action, which stimulated the continued maintenance of cumulus cells and oocyte gap junctional communication [40]. Lycopene aids in the stabilization of connexin43 mRNA [41], resulting in effective cytoplasmic maturation. The result for IGF-1 is consistent with the previous studies, which reported no significant difference between the control and IGF-1 groups in buffalo [7, 42]. This finding is due to the mechanism of action of IGF-1, which does not function through cumulus cells (CC) or obstruct the oocyte’s ability to produce an expansion factor in bovine oocytes [43]. In addition, the result of α-tocopherol is that it does not affect cumulus expansion in buffalo [44] because it is not responsible for cumulus expansion in porcine [45].
Nuclear maturation
In this study, IGF-1 and lycopene improved the oocyte nuclear maturation rate in buffalo compared with the α-tocopherol and control groups. The IGF-1 results agree with those of Pawshe
Effect of IGF-1, lycopene, and α-tocopherol supplements to IVM medium on developmental competency of mature buffalo oocytes
According to our findings, the IGF-1 and lycopene groups show significantly higher cleavage, morula, and blastocyst rates (p < 0.01) than the control and α-tocopherol groups. The results of IGF-1 are consistent with those of Ismail
Effect of IGF-1, lycopene, and α-tocopherol supplementation to IVM medium on mitochondrial function in buffalo oocytes
This work is the first to quantify mitochondrial distribution in buffalo oocytes treated with lycopene and IGF-1, addressing a significant gap in buffalo-specific mitochondrial research. There is limited information available regarding the potential benefits of antioxidant supplementation to the IVM medium on mitochondrial intensity and distribution in buffalo oocytes. The novel contribution of this study is the analysis of the mitochondrial profile through a quantitative comparison of mitochondrial intensity. Distribution categories include diffuse, semi-diffuse, and peripheral, assessed using confocal imaging for each treatment. This study found that mitochondrial intensity in the IGF-1, lycopene, and α-tocopherol groups was significantly higher (p < 0.01) than in the control group and showed the highest mitochondrial diffusion distribution, compared with the semi-diffuse and peripheral distributions across all groups. Mitochondrial distribution in equine oocytes changes dynamically during maturation. The distribution is relatively uniform in the early stages, but shifts to a heterogeneous, clustered pattern by the final MII stage. This heterogeneous distribution, characterized by a concentration of mitochondria in the inner cytoplasmic region and minimal presence in the cortex, is considered an indicator of oocyte quality and maturity. This arrangement supports the high energy demands required for fertilization and early embryonic development [67]. IGF-1 improves mitochondrial function in the current study, which agrees with the findings of Ascari
CONCLUSION
The present study demonstrated that supplementation of IVM medium with lycopene (0.2 μM) significantly enhanced cytoplasmic maturation through superior cumulus cell expansion (Grade III: 58%, GII: 28.7%; p < 0.01), nuclear maturation (87.3%; p < 0.01), and blastocyst formation compared with control, IGF-1, and α-tocopherol groups. IGF-1 (100 ng/mL) also significantly improved nuclear maturation (85.2%; p < 0.01), cleavage, morula, and blastocyst rates (p < 0.01). All three supplements increased mitochondrial intensity (p < 0.01), with IGF-1 and lycopene promoting a desirable, diffuse mitochondrial distribution, whereas α-tocopherol induced peripheral clustering and showed limited developmental benefits. Lycopene outperformed IGF-1, particularly in blastocyst yield, contradicting previous buffalo-specific findings.
These findings provide a practical, cost-effective strategy for improving oocyte quality and blastocyst production rates in buffalo
The major strength of this work is its comprehensive, multi-parameter evaluation (cytoplasmic and nuclear maturation, fertilization, embryo development to the blastocyst stage, and quantitative confocal assessment of mitochondrial intensity and distribution) conducted across three independent experiments with adequate replication. To the best of our knowledge, this is the first study to quantify mitochondrial distribution patterns in buffalo oocytes treated with lycopene, IGF-1, and α-tocopherol and to directly link these mitochondrial profiles to developmental competence.
The study was conducted exclusively
Future studies should investigate optimal dose combinations of lycopene and IGF-1, explore transcriptomic and epigenetic profiles in treated oocytes and embryos, and validate these findings through in vivo embryo transfer trials and assessments of pregnancy rates. Comparative experiments across different buffalo breeds, seasons, and oxidative stress conditions would further strengthen applicability and elucidate the underlying signaling pathways (PI3K/AKT, MAPK).
In conclusion, lycopene emerges as the most promising supplement for enhancing IVM, mitochondrial function, and embryo developmental competence in buffalo oocytes, primarily through superior antioxidant protection and maintenance of optimal mitochondrial distribution. This study provides new biological insights, reverses prior assumptions regarding IGF-1 superiority in buffalo, and offers practical strategies to overcome the inherent low efficiency of
DATA AVAILABILITY
The datasets generated during the current study are available from the corresponding author upon reasonable request.
AUTHORS’ CONTRIBUTIONS
OK: Conceptualized the study, designed the methodology, supervised the project, provided facilities, performed confocal microscope imaging, drafted the manuscript, and revised the manuscript. SE: Collected samples, conducted laboratory experiments, curated the data, analyzed the data, drafted the manuscript, and revised the manuscript. ME, SH, and NB: Analyzed the data, supervised aspects of the study, drafted the manuscript, and revised the manuscript. All authors read and approved the final manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the financial support provided by the Academy of Scientific Research and Technology (ASRT), Egypt, through its agreement with the National Natural Science Foundation of China (NSFC) under Project ID: 9154, entitled “Heat stress-induced infertility in dairy cows: Molecular basis of reduced oocyte quality and potential solution.” This work was conducted at the Embryo and Genetic Resources Conservation Bank, National Research Centre, Cairo, Egypt.
REFERENCES
- Warriach HM, McGill DM, Bush RD, Wynn PC, Chohan KR. A review of recent developments in buffalo reproduction - a review. Asian-Australas J Anim Sci 2015;28((3)):451-455. [Google Scholar] | [Crossref]
- Falchi L, Ledda S, Zedda MT. Embryo biotechnologies in sheep: achievements and new improvements. Reprod Domest Anim 2022;57((Suppl 5)):22-33. [Google Scholar] | [Crossref]
- El-Sanea AM, Abdoon ASS, Kandil OM, El-Toukhy NE, El-Maaty AMA, Ahmed HH. Effect of oxygen tension and antioxidants on the developmental competence of buffalo oocytes cultured
in vitro . Vet World 2021;14((1)):78-84. [Google Scholar] | [Crossref] - Almeeni IM. The tissue culture media supplement to improve
in vitro embryos production in ewes: a review. Diyala J Vet Sci 2023;1((4)):136-152. [Google Scholar] | [Crossref] - Lu F, Luo T, Sun H, Li N, Liu X, Meng L, Jiang J, Shi D. Effects of insulin-like growth factor 1 (IGF-1) on the development and apoptosis of preimplantation buffalo (
Bubalus bubalis ) embryo. Reprod Fertil Dev 2012;25:215-216. [Google Scholar] | [Crossref] - Singh N, Dhaliwal G, Brar P, Singhal S, Malik V.
In vitro maturation of buffalo oocytes in serum-free media supplemented with growth factors. Haryana Vet 2021;60((SI)):1-4. [Google Scholar] | [Crossref] - Ismail EA, Elsayed MAI, Hemeida NA, Abdoon ASS, Kandil OM. Effect of insulin-like growth factor-I (IGF-1) on oocyte competence and embryo development of buffaloes (
Bubalus bubalis ). Int J ChemTech Res 2016;9((5)):738-748. [Google Scholar] | [Crossref] - Meiyu QI, Roth Z, LIU Di. Insulin-like growth factor-I (IGF-I) in the reproductive system of female bovine. J Northeast Agric Univ (English Ed) 2011;18((4)):84-87. [Google Scholar] | [Crossref]
- Leh HE, Lee LK. Lycopene: a potent antioxidant for the amelioration of type II diabetes mellitus. Molecules 2022;27((7)):2335. [Google Scholar] | [Crossref]
- Black HS, Boehm F, Edge R, Truscott TG. The benefits and risks of certain dietary carotenoids that exhibit both anti- and pro-oxidative mechanisms - a comprehensive review. Antioxidants (Basel) 2020;9((3)):264. [Google Scholar] | [Crossref]
- Chowdhury MMR, Mesalam A, Khan I, Joo MD, Lee KL, Xu L, Afrin F, Kong IK. Improved developmental competence in embryos treated with lycopene during
in vitro culture system. Mol Reprod Dev 2018;85((1)):46-61. [Google Scholar] | [Crossref] - Sidi S, Residewati G. Antioxidant effect of lycopene on
in vitro maturation and fertilization of bovine oocytes. FUDMA J Agric Agric Technol 2024;10((4)):82-87. [Google Scholar] | [Crossref] - Sidi S, Pascottini OB, Angel-Velez D, Azari-Dolatabad N, Pavani KC, Residiwati G, Meese T, Van Nieuwerburgh F, Kambai Bawa E, Alikidon Voh A, Olusegun Ayo J, Van Soom A. Lycopene supplementation to serum-free maturation medium improves
in vitro bovine embryo development and quality and modulates embryonic transcriptomic profile. Antioxidants 2022;11((2)):344. [Google Scholar] | [Crossref] - Residiwati G, Azari-Dolatabad N, Tuska HSA, Sidi S, Van Damme P, Benedetti C, Montoro AF, Luceno NL, Budiono, Pavani KC, Opsomer G, Van Soom A, Bogado Pascottini O. Effect of lycopene supplementation to bovine oocytes exposed to heat shock during
in vitro maturation. Theriogenology 2021;173:48-55. [Google Scholar] | [Crossref] - Olson SE, Seidel GE. Culture of
in vitro -produced bovine embryos with vitamin E improves developmentin vitro and after transfer to recipients. Biol Reprod 2000;62((2)):248-252. [Google Scholar] | [Crossref] - Thiyagarajan B, Valivittan K. Ameliorating effect of vitamin E on
in vitro development of preimplantation buffalo embryos. J Assist Reprod Genet 2009;26((4)):217-225. [Google Scholar] | [Crossref] - Báez F, Gómez B, de Brun V, Rodríguez-Osorio N, Viñoles C. Effect of ethanol on parthenogenetic activation and α-tocopherol supplementation during
in vitro maturation on developmental competence of summer-collected bovine oocytes. Curr Issues Mol Biol 2021;43((3)):2253-2265. [Google Scholar] | [Crossref] - McCluskey S, Hall M, Stanton C, Devery R. Alpha-tocopherol inhibits oxidative stress induced by cholestanetriol and 25-hydroxycholesterol in porcine ovarian granulosa cells. Mol Cell Biochem 1999;194((1-2)):217-225. [Google Scholar] | [Crossref]
- Tareq KMA, Akter QS, Khandoker MAMY, Tsujii H. Selenium and vitamin E improve the
in vitro maturation, fertilization and culture to blastocyst of porcine oocytes. J Reprod Dev 2012;58((6)):621-628. [Google Scholar] | [Crossref] - Natarajan R, Shankar MB, Munuswamy D. Effect of α-tocopherol supplementation on
in vitro maturation of sheep oocytes andin vitro development of preimplantation sheep embryos to the blastocyst stage. J Assist Reprod Genet 2010;27:483-490. [Google Scholar] | [Crossref] - Van Metre DC, Callan RJ. Selenium and vitamin E. Vet Clin North Am Food Anim Pract 2001;17((2)):373-402. [Google Scholar] | [Crossref]
- Dalvit G, Llanes SP, Descalzo A, Insani M, Beconi M, Cetica P. Effect of alpha-tocopherol and ascorbic acid on bovine oocyte
in vitro maturation. Reprod Domest Anim 2005;40((2)):93-97. [Google Scholar] | [Crossref] - Arias-Álvarez M, García-García RM, López-Tello J, Rebollar PG, Gutiérrez-Adán A, Lorenzo PL. α-Tocopherol modifies the expression of genes related to oxidative stress and apoptosis during
in vitro maturation and enhances the developmental competence of rabbit oocytes. Reprod Fertil Dev 2018;30((12)):1728-1738. [Google Scholar] | [Crossref] - Kirillova A, Smitz JEJ, Sukhikh GT, Mazunin I. The role of mitochondria in oocyte maturation. Cells 2021;10((9)):2484. [Google Scholar] | [Crossref]
- Biswas S, Ghosh S, Maitra S. Role of insulin-like growth factor 1 (IGF1) in the regulation of mitochondrial bioenergetics in zebrafish oocytes: lessons from
in vivo andin vitro investigations. Front Cell Dev Biol 2023;11:1202693. [Google Scholar] | [Crossref] - Kang HG, Lee S, Jeong PS, Kim MJ, Park SH, Joo YE, Park SH, Song BS, Kim SU, Kim MK, Sim BW. Lycopene improves
in vitro development of porcine embryos by reducing oxidative stress and apoptosis. Antioxidants 2021;10((2)):230. [Google Scholar] | [Crossref] - Lauridsen C, Jensen SK. α-Tocopherol incorporation in mitochondria and microsomes upon supranutritional vitamin E supplementation. Genes Nutr 2012;7:475-482. [Google Scholar] | [Crossref]
- Feng Z, Shi J, Ren J, Luo L, Liu D, Guo Y, Sun B, Liu G, Deng M, Li Y. Mitochondria-targeted antioxidant MitoQ improves
in vitro maturation and subsequent embryonic development from culled cows. Animals 2024;14((20)):2929. [Google Scholar] | [Crossref] - Zhang H, Li C, Wen D, Li R, Lu S, Xu R, Tang Y, Sun Y, Zhao X, Pan M, Ma B. Melatonin improves the quality of maternally aged oocytes by maintaining intercellular communication and antioxidant metabolite supply. Redox Biol 2022;49:102215. [Google Scholar] | [Crossref]
- Lee IW, Tazehkand AP, Sha Z-Y, Adhikari D, Carroll J. An aggregated mitochondrial distribution in preimplantation embryos disrupts nuclear morphology, function, and developmental potential. Proc Natl Acad Sci U S A 2024;121((27)):e2317316121. [Google Scholar] | [Crossref]
- Murphy MP, Smith RAJ. Targeting antioxidants to mitochondria by conjugation to lipophilic cations. Annu Rev Pharmacol Toxicol 2007;47:629-656. [Google Scholar] | [Crossref]
- Hao X, Zhao J, Rodriguez-Wallberg KA. Comprehensive atlas of mitochondrial distribution and dynamics during oocyte maturation in mouse models. Biomark Res 2024;12:125. [Google Scholar] | [Crossref]
- Gałęska E, Kowalczyk A, Wrzecińska M, García MC, Czerniawska-Piątkowska E, Gwoździewicz S, Witkiewicz W, Dobrzański Z. The importance of mitochondrial processes in the maturation and acquisition of competences of oocytes and embryo culture. Int J Mol Sci 2025;26((9)):4098. [Google Scholar] | [Crossref]
- Leibfried-Rutledge ML, Critser ES, Eyestone WH, Northey DL, First NL. Development potential of bovine oocytes matured
in vitro orin vivo . Biol Reprod 1987;36((2)):376-383. [Google Scholar] | [Crossref] - Kandil OM, Alhallag KA, Shawky SM, Abu-alya IS, Kandeel SM, Ismail EA, Fathalla SI. Effect of zinc and nano zinc on developmental competence of buffalo oocytes. J Adv Vet Res 2023;13((3)):474-478. [Google Scholar] | [Crossref]
- Kandil OM, Abdoon ASS, Murakami M, Otoi T, Suzuki T. New technique, using a portable CO₂incubator, for the production of
in vitro fertilized Egyptian buffalo embryos. J Reprod Dev 1999;45((5)):315-320. [Google Scholar] | [Crossref] - Bleil JD, Wassarman PM. Synthesis of zona pellucida proteins by denuded and follicle-enclosed mouse oocytes during culture
in vitro . Proc Natl Acad Sci USA 1980;77((2)):1029-1033. [Google Scholar] | [Crossref] - Ismail EA, Elsayed MAI, Hemeida NA, Abdoon ASS, Kandil OM. Effect of different cryoprotectant agents on mitochondrial distribution and developmental competence in buffalo oocyte (
Bubalus bubalis ). Egypt J Chem 2022;65((1)):539-553. [Google Scholar] | [Crossref] - Beath A, Jones MP. Guided by the research design: choosing the right statistical test. Med J Aust 2018;208((4)):163-165. [Google Scholar] | [Crossref]
- Watanabe H, Okawara S, Bhuiyan MMU, Fukui Y. Effect of lycopene on cytoplasmic maturation of porcine oocytes
in vitro . Reprod Domest Anim 2010;45((5)):838-845. [Google Scholar] | [Crossref] - Heber D, Lu QY. Overview of mechanisms of action of lycopene. Exp Biol Med (Maywood) 2002;227((10)):920-923. [Google Scholar] | [Crossref]
- Kumar D, Purohit GN. Effect of epidermal and insulin-like growth factor-1 on cumulus expansion, nuclear maturation and fertilization of buffalo cumulus oocyte complexes in simple serum free media DMEM and Ham's F-10. Vet Arh 2004;74((1)):13-25. [Google Scholar] | [Crossref]
- Lorenzo PL, Illera MJ, Illera JC, Illera M. Enhancement of cumulus expansion and nuclear maturation during bovine oocyte maturation
in vitro by the addition of epidermal growth factor and insulin-like growth factor I. J Reprod Fertil 1994;101((3)):697-701. [Google Scholar] | [Crossref] - Azam A, Ejaz R, Qadeer S, Irum S, Ul-Husna A, Ullah S, Shahzad Q, Akhtar T, Akhter S. Synergistic impact of α-linolenic acid and α-tocopherol on
in vitro maturation and culture of buffalo oocytes. Brazilian J Biol 2024;84:e253514. [Google Scholar] | [Crossref] - Tao Y, Zhou B, Xia G, Wang F, Wu Z, Fu M. Exposure to L-ascorbic acid or α-tocopherol facilitates the development of porcine denuded oocytes from metaphase I to metaphase II and prevents cumulus cells from fragmentation. Reprod Domest Anim 2004;39((1)):52-57. [Google Scholar] | [Crossref]
- Pawshe CH, Appa Rao KBC, Totey SM. Effect of insulin-like growth factor I and its interaction with gonadotropins on
in vitro maturation and embryonic development, cell proliferation, and biosynthetic activity of cumulus-oocyte complexes and granulosa cells in buffalo. Mol Reprod Dev 1998;49((3)):277-285. [Google Scholar] | [Crossref] - Hernandez-Fonseca HJ, Sirisathien S, Bosch P, Cho HS, Lott JD, Hawkins LL, Hollett RB, Coley SL, Brackett BG. Offspring resulting from direct transfer of cryopreserved bovine embryos produced
in vitro in chemically defined media. Anim Reprod Sci 2002;69((3-4)):151-158. [Google Scholar] | [Crossref] - Quetglas MD, Coelho LA, Garcia JM, Oliveira Filho EB, Esper CR. Effect of insulin-like growth factor-1 during
in vitro oocyte maturation andin vitro culture of bovine embryos. Arq Bras Med Vet Zootec 2001;53((2)):207-211. [Google Scholar] | [Crossref] - Lee ES, Fukui Y. Effect of various growth factors in a defined culture medium on
in vitro development of bovine embryos matured and fertilizedin vitro . Theriogenology 1995;44((1)):71-83. [Google Scholar] | [Crossref] - Mazerbourg S, Bondy CA, Zhou J, Monget P. The insulin-like growth factor system: a key determinant role in the growth and selection of ovarian follicles?A comparative species study. Reprod Domest Anim 2003;38((4)):247-258. [Google Scholar] | [Crossref]
- Bezerra MES, Barberino RS, Menezes VG, Gouveia BB, Macedo TJS, Santos JMS, Monte APO, Barros VRP, Matos MHT. Insulin-like growth factor-1 (IGF-1) promotes primordial follicle growth and reduces DNA fragmentation through the phosphatidylinositol 3-kinase/protein kinase B (PI3K/AKT) signalling pathway. Reprod Fertil Dev 2018;30((11)):1503-1513. [Google Scholar] | [Crossref]
- Hainaut P, Giorgetti S, Kowalski A, Ballotti R, Van Obberghen E. Antibodies to phosphotyrosine injected in
Xenopus laevis oocytes modulate maturation induced by insulin/IGF-I. Exp Cell Res 1991;195((1)):129-136. [Google Scholar] | [Crossref] - Tatemoto H, Terada T. Activation of p34cdc2 kinase around the meiotic resumption in bovine oocytes cultured
in vitro . Theriogenology 1996;45((2)):427-437. [Google Scholar] | [Crossref] - Sakaguchi M, Dominko T, Yamauchi N, Leibfried-Rutledge ML, Nagai T, First NL. Possible mechanism for acceleration of meiotic progression of bovine follicular oocytes by growth factors
in vitro . Reproduction 2002;123((1)):135-142. [Google Scholar] | [Crossref] - Hayes E, Winston N, Stocco C. Molecular crosstalk between insulin-like growth factors and follicle-stimulating hormone in the regulation of granulosa cell function. Reprod Med Biol 2024;23((1)):e12575. [Google Scholar] | [Crossref]
- Spicer LJ, Aad PY. Insulin-like growth factor (IGF) 2 stimulates steroidogenesis and mitosis of bovine granulosa cells through the IGF1 receptor: role of follicle-stimulating hormone and IGF2 receptor. Biol Reprod 2007;77((1)):18-27. [Google Scholar] | [Crossref]
- Araujo MS, Guastali MD, Paulini F, Silva AN, Tsunemi MH, Fontes PK, Castilho ACS, Landim-Alvarenga FC. Molecular and cellular effects of insulin-like growth factor-1 and LongR3-IGF-1 on
in vitro maturation of bovine oocytes: comparative study. Growth Horm IGF Res 2020;55:101357. [Google Scholar] | [Crossref] - Chowdhury MMR, Choi BH, Khan I, Lee KL, Mesalam A, Song SH, Xu L, Joo MD, Afrin F, Kong IK. Supplementation of lycopene in maturation media improves bovine embryo quality
in vitro . Theriogenology 2017;103:173-184. [Google Scholar] | [Crossref] - Bury L, Coelho PA, Glover DM. From meiosis to mitosis: the astonishing flexibility of cell division mechanisms in early mammalian development. Curr Top Dev Biol 2016;120:1-42. [Google Scholar] | [Crossref]
- Adeldust H, Zeinoaldini S, Kohram H, Roudbar MA, Joupari MD.
In vitro maturation of ovine oocyte in a modified granulosa cells co-culture system and alpha-tocopherol supplementation: effects on nuclear maturation and cleavage. J Anim Sci Technol 2015;57:27. [Google Scholar] | [Crossref] - Byrne AT, Southgate J, Brison DR, Leese HJ. Regulation of apoptosis in the bovine blastocyst by insulin and the insulin-like growth factor (IGF) superfamily. Mol Reprod Dev 2002;62((4)):489-495. [Google Scholar] | [Crossref]
- Block J, Wrenzycki C, Niemann H, Herrmann D, Hansen PJ. Effects of insulin-like growth factor-1 on cellular and molecular characteristics of bovine blastocysts produced
in vitro . Mol Reprod Dev 2008;75((5)):895-903. [Google Scholar] | [Crossref] - Yang R fen, Xiong X rong, Zi X dong. Effect of cysteine, insulin-like growth factor-1 and epidermis growth factor during
in vitro oocyte maturation andin vitro culture of yak-cattle crossbred embryos. J Appl Anim Res 2019;47((1)):463-466. [Google Scholar] | [Crossref] - Fernandez-Gonzalez L, Kozhevnikova V, Brusentsev E, Jänsch S, Amstislavsky S, Jewgenow K. IGF-1 medium supplementation improves singly cultured cat oocyte maturation and embryo development
in vitro . Animals 2021;11((7)):1909. [Google Scholar] | [Crossref] - Singhal S, Prasad S, Singh B, Prasad JK, Gupta HP. Effect of including growth factors and antioxidants in maturation medium used for
in vitro culture of buffalo oocytes recoveredin vivo . Anim Reprod Sci 2009;113((1-4)):44-50. [Google Scholar] | [Crossref] - Quirk SM, Harman RM, Cowan RG. Regulation of Fas antigen (Fas, CD95)-mediated apoptosis of bovine granulosa cells by serum and growth factors. Biol Reprod 2000;63((5)):1278-1284. [Google Scholar] | [Crossref]
- Luis-Calero M, Fernández-Hernández P, Ortiz-Rodríguez JM, Muñoz-García CC, Jardin I, Macías-García B, González-Fernández L. Description of a new quantitative method to assess mitochondrial distribution pattern in mature equine oocytes. Vet Res Commun 2024;48((3)):1867-1871. [Google Scholar] | [Crossref]
- Ascari IJ, Alves NG, Jasmin J, Lima RR, Quintão CCR, Oberlender G, Moraes EA, Camargo LSA. Addition of insulin-like growth factor I to the maturation medium of bovine oocytes subjected to heat shock: effects on the production of reactive oxygen species, mitochondrial activity and oocyte competence. Domest Anim Endocrinol 2017;60:50-60. [Google Scholar] | [Crossref]
- Ispada J, Lima RS, Risolia P, Assumpção MEOA, Visintin JA, Paula-Lopes FF. Insulin-like growth factor-1 exerts a thermoprotective role on mitochondrial function of bovine oocytes exposed to heat shock. Reprod Fertil Dev 2011;24((1)):209. [Google Scholar] | [Crossref]
- Poudel SB, Dixit M, Neginskaya M, Nagaraj K. Effects of GH/IGF on the aging mitochondria. Cells 2020;9((6)):1384. [Google Scholar] | [Crossref]
- Shamas-Din A, Kale J, Leber B, Andrews DW. Mechanisms of action of Bcl-2 family proteins. Cold Spring Harb Perspect Biol 2013;5((4)):a008714. [Google Scholar] | [Crossref]
- Danial NN, Korsmeyer SJ. Cell death: critical control points. Cell 2004;116((2)):205-219. [Google Scholar] | [Crossref]
- Fernández-Montoro A, Angel-Velez D, Cava-Cami B, Pascottini OB, Pavani KC, Smits K, Van Soom A. How to beat the bull: lycopene as a tool to improve
in vitro fertilization efficiency in bulls with high polyspermy. Reprod Biol 2024;24((2)):100888. [Google Scholar] | [Crossref] - Napolitano G, Fasciolo G, Muscari Tomajoli MT, Venditti P. Changes in the mitochondria in the aging process—can α-tocopherol affect them?. Int J Mol Sci 2023;24((15)):12453. [Google Scholar] | [Crossref]