


ABSTRACT
Background and Aim: The
Materials and Methods: A total of 1,160 healthy LCGs (89 bucks and 1,071 does, 2–4 years of age) from the same nucleus herd were included. The C31799T locus in
Results: The TT genotype at the
Conclusion: This is the first study to establish
Keywords: cashmere fineness, Cashmere goat, Cashmere production traits, cashmere yield, GMPS gene, Liaoning Cashmere goat, marker-assisted selection, SRC gene.
INTRODUCTION
Cashmere, hailed as the “gem of fibers” [1], is a precious textile material, and cashmere goat farming constitutes a vital economic foundation for many regions. The
Sex hormones represent the core regulators of hair follicle development and cycling. The
Although high-throughput sequencing technologies have accelerated the identification of genetic markers linked to fiber traits across diverse cashmere goat populations, the roles of polymorphisms in the
The present study was therefore designed to address this gap through a comprehensive evaluation of the sex-specific effects of selected single-nucleotide polymorphisms (SNPs) in
MATERIALS AND METHODS
Ethical approval
All procedures involving live animals in this study were reviewed and approved by the Laboratory Animal Management and Use Ethics Committee of Shenyang Agricultural University (Approval No. 20240513). The experiment was conducted in full compliance with the institutional guidelines for the care and use of experimental animals and the relevant national regulations on animal welfare in China (including the Regulations on the Administration of Laboratory Animals and the Guidelines for the Ethical Review of Laboratory Animal Welfare).
The study was performed at the LCG Breeding Center, Liaoyang City, Liaoning Province, China. All blood sample collections were carried out under the direct supervision of a licensed veterinarian to ensure minimal stress and discomfort to the animals. No invasive surgical procedures were performed, and all handling was conducted according to the principles of the 3Rs (Replacement, Reduction, and Refinement). The animals remained under standard farm management conditions throughout the study, and no additional pain, suffering, or distress beyond routine husbandry practices was imposed.
Written informed consent for the use of the animals was obtained from the farm management prior to the commencement of the study. The authors confirm that the research adheres to the ARRIVE 2.0 guidelines (Animal Research: Reporting of
Study period and location
This study was conducted from April to August 2025 at the LCG Breeding Center, Liaoyang City, Liaoning Province, China (approximate coordinates: 41.24° N, 123.14° E).
Animals, inclusion criteria, and management conditions
A total of 1,160 clinically healthy LCG were enrolled in the study (89 bucks and 1,071 does; age range 2–4 years). All animals belonged to the same nucleus breeding herd and were therefore at a comparable production stage. Inclusion criteria were as follows: clinically healthy appearance, absence of major parasitic infestations, and no history of antibiotic treatment within the preceding 4 weeks. Animals were maintained under semi-intensive management conditions with a stocking density of approximately 15–20 goats per pen. They had
Phenotypic trait recording and definitions
All phenotypic measurements were performed by the same fixed team of five trained technicians to ensure maximum consistency and to minimize inter-observer variation. Detailed definitions, measurement timing, units, and protocols for each recorded trait are presented in Supplementary Table S1. The main instruments and corresponding calibration procedures are described below.
Measurement of cashmere production traits and instrument calibration
CF (fiber diameter) and staple length were determined using the portable all-weather CF and length analyzer (Model: OFDA2000, BSC Electronics, Ardross, Australia). The instrument was calibrated weekly before each measurement session using a certified standard cashmere bundle (nominal fineness 15.0 ± 0.1 μm) according to the Chinese national standard Test Method for Cashmere Fiber (GB/T 18267-2019). The calibration deviation was maintained at ≤ 0.2 μm throughout the study. Collected cashmere samples were evenly spread on a dedicated sample slide, the slide was inserted into the detection chamber, and the “Start Test” function was activated to record fiber diameter and length.
Measurement of body size traits and instrument calibration
Body conformation measurements were obtained using an intelligent three-dimensional optical body measurement system (3D Optical Body Scanner, Model: VITUS Smart, Human Solutions GmbH, Kaiserslautern, Germany). Animals were gently guided into the measurement zone where the system automatically initiated multi-angle scanning. Recorded parameters included: body height, withers height, body length, chest depth, chest width, hip width, heart girth, cannon circumference, and pin bone height. The instrument was calibrated monthly using a standard reference mannequin, achieving a measurement error of ≤ 0.5 cm.
Measurement of milk composition and instrument calibration
Milk composition was analyzed using the MilkoScan FT120 milk analyzer (Foss Electric A/S, Hillerød, Denmark). The following parameters were determined: milk fat, milk protein, lactose, urea nitrogen, non-fat solids, total solids, conductivity, and health index. Daily calibration was performed before each analysis session using Foss calibration milk (Product No.: 8011-0001, Foss Electric) according to ISO 9622:2013. Calibration error for the major components (fat, protein, lactose) was maintained at ≤ 0.05%. The built-in conductivity sensor was simultaneously calibrated (measurement range 0–20 mS/cm; accuracy ±0.05 mS/cm).
Blood collection and storage
Whole blood was collected from the jugular vein into commercial ethylenediaminetetraacetic acid vacuum blood collection tubes (Sangon Biotech, Shanghai, China). Immediately after collection, tubes were placed on ice, transported to the laboratory within 4 hours, and stored at –20 °C until DNA extraction (maximum storage duration 2 weeks).
Extraction and quality assessment of genomic DNA
Genomic DNA was extracted from 200 µL of anticoagulated whole blood using the Ezup column animal genomic DNA extraction kit (Cat. No. B518253-0100, Sangon Biotech, Shanghai, China) following the manufacturer’s protocol. In brief: Proteinase K and lysis buffer (Buffer DL) were added, followed by incubation at 56°C for 10 min. After addition of absolute ethanol and thorough mixing, the lysate was transferred to the adsorption column. The column was washed sequentially with GW solution and Wash solution (two washes each), followed by a high-speed centrifugation step to remove residual Wash buffer. DNA was finally eluted with Capillary Electrophoresis Buffer. DNA purity and concentration were assessed using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Samples were considered acceptable when the A260/A280 ratio ranged between 1.8 and 2.0 and the concentration exceeded 50 ng/µL. Qualified DNA samples were stored at –20°C until further use.
SNP discovery and sequence alignment
The target SNP loci C31799T (
Primer design and PCR amplification
Gene-specific primers flanking the target intronic regions of
Table 1. Amplification and genotyping primers for target fragments of
| Gene | Sense primer (5′→3′) | Anti-sense primer (5′→3′) | Tm (°C) | Fragment size (bp) | Genomic region |
|---|---|---|---|---|---|
|
| ATCTGCTGTGGCATACTGA | CACTGATGAGCCTGTGAAA | 48.4 | 754 | 111285624–111357129 |
|
| TCATCTGTGCTTGACCCTA | CCCTAAGACCAAATAACCC | 52 | 637 | 65852275–65906216 |
PCR was performed in a total volume of 50 µL containing:
25 µL of 2× SanTaq PCR Mix (Sangon Biotech, Shanghai, China) 1 µL DNA template (~50 ng) 2 µL of each primer (final concentration 0.4 µM each) nuclease-free water to final volume Thermal cycling conditions were as follows: Initial denaturation 94°C / 5 min 35 cycles of: 94°C / 30 s 48.4°C ( 72°C / 30 s Final extension 72°C / 10 min
PCR product verification and Sanger sequencing
Amplification products were separated on 1.5% (w/v) agarose gels prepared in 1× TAE buffer and stained with GelRed nucleic acid stain. Electrophoresis was conducted at 130 V (constant power 180 W) for 20 min. A 100 bp DNA ladder (Thermo Fisher Scientific) was included for size determination. Amplicons showing a single band of the expected size were purified using the SanPrep Column PCR Product Purification Kit (Sangon Biotech, Shanghai, China) according to the manufacturer’s instructions. Purified products were submitted for bidirectional Sanger sequencing (Sangon Biotech Co., Ltd., Shanghai, China).
Genotyping and quality control
Genotypes were manually called from bidirectional sequencing chromatograms using Chromas software version 2.6.6. A heterozygous call was accepted only when the secondary peak height reached at least 35% of the primary peak height. All chromatograms were independently scored by two researchers. Samples exhibiting poor sequence quality or a genotype call missing rate >5% were excluded. To assess repeatability, the complete workflow (PCR → sequencing → genotyping) was repeated for 10% of randomly selected samples, resulting in 100% genotype concordance.
Haplotype inference
Haplotype phase was inferred using the SHEsis online platform with default parameters. To allow detection of sex-specific haplotype effects, haplotype reconstruction was performed separately for bucks and does. Rare haplotype combinations (population frequency <1%) were collapsed into a single “other” category for subsequent association analyses.
Statistical analysis
Data distribution assumptions
Normality of continuous trait distributions was confirmed using the Shapiro–Wilk test (p > 0.05). Homogeneity of variance across genotype groups was verified with Levene’s test (p > 0.05). Both assumptions were satisfied, permitting the use of parametric statistical procedures.
Genotype–trait association analysis
Associations between genotypes and phenotypic traits were initially evaluated by one-way analysis of variance (ANOVA). Comprehensive analyses were performed using the following general linear mixed model:
Yijkl = μ + gj + bk + sl + eijkl
where Yijkl = observed phenotypic value μ = overall mean gj = fixed effect of genotype bk = fixed effect of age class sl = fixed effect of sex eijkl = random residual error
Multiple comparisons
When ANOVA indicated significant effects, pairwise comparisons were conducted using Duncan’s multiple range test. Results are presented as mean ± standard error of the mean (SEM). Levels of significance: p > 0.05 not significant; p < 0.05 significant (different lowercase letters); p < 0.01 highly significant (different uppercase letters).
Correlation, path, and regression analyses
Pearson correlation coefficients were calculated for pairwise trait relationships. Path analysis was performed to partition direct and indirect effects. Stepwise multiple regression was conducted (entry criterion p < 0.05; removal criterion p > 0.10). Multicollinearity was assessed using the variance inflation factor (VIF); all final model predictors had VIF < 3.
Software and significance threshold
Statistical analyses were performed using SPSS version 27.0 (ANOVA, correlation, regression), Microsoft Excel (genetic parameter calculations), SHEsis (haplotype inference), Chromas v. 2.6.6 (chromatogram viewing), and DNAMAN v. 9.0 (sequence alignment). The significance threshold was set at p < 0.05; p < 0.01 was considered highly significant. All continuous data are reported as mean ± SEM.
RESULTS
PCR amplification of GMPS and SRC
Figure 1 shows the agarose gel electrophoresis results of PCR products for the target loci. (Left panel) A single specific band of 754 bp corresponding to the
Figure 1. Polymerase chain reaction amplification of (A)
Identification of SNP
Figure 2 presents representative bidirectional sequencing chromatograms of the target polymorphic sites. (Left)
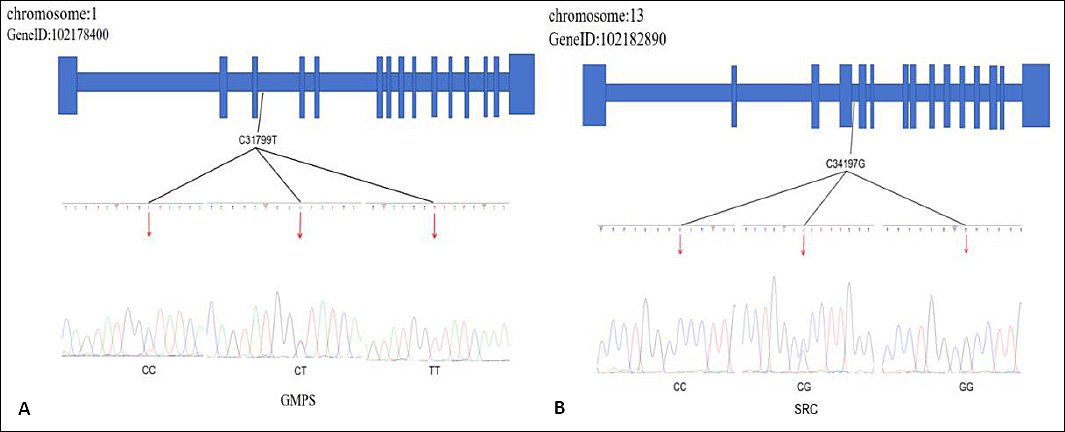
Figure 2. Identification of SNPs in (A)
Genetic polymorphisms of the GMPS and SRC genes in the LCG
Genotype frequencies, allele frequencies, polymorphism information content (PIC), expected heterozygosity (He), effective number of alleles (Ne), and Hardy–Weinberg equilibrium χ² test results at the two loci are presented in Table 2. The T allele at
Table 2. The genetic structure of the population at the polymorphic sites of the
| Gene | Loci | Gender | Genotype frequency | Allele frequency | PIC | He | Ne | χ² | p-value |
|---|---|---|---|---|---|---|---|---|---|
|
| C31799T | Buck | CC 0.07, CT 0.23, TT 0.70 | T 0.81, C 0.19 | 0.26 | 0.30 | 1.43 | 7.51 | 0.01 |
| Doe | CC 0.03, CT 0.38, TT 0.59 | T 0.78, C 0.22 | 0.28 | 0.34 | 1.52 | 12.44 | 0.0004 | ||
|
| C34197G | Buck | CC 0.59, CG 0.41, GG 0.00 | C 0.80, G 0.20 | 0.27 | 0.33 | 1.48 | 7.27 | 0.007 |
| Doe | CC 0.69, CG 0.24, GG 0.07 | C 0.81, G 0.19 | 0.26 | 0.31 | 1.45 | 54.25 | 1.76×10⁻¹³ |
Effect of gene substitution
Additive and dominance effects estimated for the two loci are summarized in Table 3. Substitution of the C allele by the T allele at
Table 3. Gene substitution effect analysis of single-nucleotide polymorphisms in
| Gene | Loci | Gender | Dominant effect | Additive effect | B gene average effect | A gene average effect | Average effect of B instead of A |
|---|---|---|---|---|---|---|---|
|
| C31799T | Buck | –18.00 | 34.00 | 8.40 | –36.94 | 45.33 |
| Doe | 70.00 | 273.00 | 51.28 | –182.44 | 233.72 | ||
|
| C34197G | Buck | 12.50 | –32.50 | –19.98 | 5.14 | –25.11 |
| Doe | –140.00 | –300.00 | –311.70 | 74.37 | –386.07 |
Effects of GMPS and SRC genes on cashmere production traits in LCG
Genotype means ± standard error and significance levels for cashmere production traits are presented in Table 4. In bucks, the
Table 4. Cashmere production performance of the
| Gender | Gene | Loci | Genotype (n/total) | Shearing quantity (g) | Fineness (μm) | Length (cm) | Coefficient of length variation (%) | Curl number | Short fiber rate (%) | Cashmere yield rate (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| Buck |
| C31799T | CC (6/78) | 1850.00 ± 67.08 | 16.75 ± 0.37 | 118.80 ± 16.10Aa | 46.60 ± 1.96 | 9.05 ± 0.16 | 0.09 ± 0.02 | 0.73 ± 0.01 |
| CT (18/78) | 2066.67 ± 75.30 | 16.94 ± 0.14 | 95.47 ± 4.91ABb | 49.95 ± 1.93 | 6.88 ± 0.77 | 0.13 ± 0.02 | 0.69 ± 0.02 | |||
| TT (54/78) | 1944.44 ± 51.03 | 16.37 ± 0.17 | 90.76 ± 2.97Bb | 57.31 ± 1.93 | 7.15 ± 0.54 | 0.21 ± 0.02 | 0.78 ± 0.02 | |||
| Doe |
| C31799T | CC (14/1022) | 2350.00 ± 12.65Aa | 18.09 ± 12.68Aa | 61.60 ± 1.34Bb | 25.39 ± 12.68Bb | 5.24 ± 12.69 | 0.21 ± 12.68 | 0.54 ± 12.68Bb |
| CT (448/1022) | 1753.13 ± 12.71Bb | 16.72 ± 12.71Bb | 100.6 ± 1.53Aa | 53.57 ± 12.71Aa | 3.13 ± 12.71 | 0.17 ± 12.71 | 0.74 ± 12.71Aa | |||
| TT (560/1022) | 1798.75 ± 12.62Bb | 16.52 ± 12.62Bb | 97.73 ± 1.26Aa | 54.77 ± 12.62Aa | 8.75 ± 12.62 | 0.16 ± 12.62 | 0.76 ± 12.62Aa | |||
| Buck |
| C34197G | CC (44/80) | 2000.00 ± 36.00 | 16.84 ± 0.15 | 91.35 ± 2.61 | 54.96 ± 1.96 | 8.02 ± 0.37 | 0.13 ± 0.02ABab | 0.70 ± 0.02 |
| CG (36/80) | 2127.78 ± 52.24 | 16.58 ± 0.18 | 98.36 ± 4.70 | 56.77 ± 1.86 | 6.98 ± 0.58 | 0.27 ± 0.03Aa | 0.79 ± 0.03 | |||
| Doe |
| C34197G | CC (686/1022) | 1828.57 ± 12.01Aa | 16.81 ± 0.05Aa | 96.77 ± 1.21 | 53.85 ± 0.54 | 6.43 ± 0.15Bb | 0.18 ± 0.01Aa | 0.73 ± 0.01Bb |
| CG (252/1022) | 1734.86 ± 15.04Bb | 16.18 ± 0.07Bb | 97.79 ± 1.74 | 52.34 ± 0.67 | 7.58 ± 0.24Aa | 0.12 ± 0.01Ba | 0.79 ± 0.01Aa | |||
| GG (84/1022) | 1816.67 ± 38.51ABa | 16.42 ± 0.14Bb | 99.07 ± 3.45 | 53.79 ± 1.58 | 5.70 ± 0.45Bb | 0.20 ± 0.01Aa | 0.78 ± 0.01Aa |
Different lowercase letters indicate significant differences (p < 0.05); different uppercase letters indicate highly significant differences (p < 0.01). Sample sizes were limited for some genotype groups (e.g.,
Effects of
Table 5. Analysis of the body size traits of the
| Gender | Gene | Loci | Genotype (n/total) | Body height (cm) | Height at the sacrum (cm) | Body length (cm) | Chest depth (cm) | Chest width (cm) | Waist width (cm) | Chest circumference (cm) | Tube circumference (cm) | Waist height (cm) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Doe |
| C31799T | CC (32/1064) | 64.25 ± 0.45 | 67.50 ± 0.53Aa | 71.75 ± 1.11Bb | 33.80 ± 0.27Aa | 25.00 ± 0.22Aa | 25.88 ± 0.49Aa | 100.25 ± 1.31ABab | 10.25 ± 0.19Aa | 65.00 ± 0.77 |
| CT (400/1064) | 63.69 ± 0.19 | 65.09 ± 0.19Bb | 77.65 ± 0.33Aa | 32.31 ± 0.14Bb | 23.24 ± 0.22Bb | 22.52 ± 0.23Bb | 97.87 ± 0.40Bb | 9.48 ± 0.05Bb | 63.97 ± 0.22 | |||
| TT (632/1064) | 64.06 ± 0.13 | 65.56 ± 0.13Bb | 79.10 ± 0.24Aa | 32.52 ± 0.11Bb | 23.99 ± 0.15ABab | 23.48 ± 0.18Bb | 101.71 ± 0.40Aa | 9.37 ± 0.04Bb | 64.56 ± 0.19 | |||
| Buck |
| C31799T | CC (6/81) | 75.50 ± 0.19 | 80.25 ± 1.04Aa | 89.50 ± 0.57 | 34.75 ± 0.28 | 25.25 ± 1.04ab | 19.00 ± 1.51 | 104.75 ± 1.80ABab | 12.50 ± 0.23 | 76.00 ± 0.38Aa |
| CT (18/81) | 75.50 ± 0.82 | 72.42 ± 0.54Bb | 86.83 ± 1.42 | 36.58 ± 0.30 | 23.50 ± 0.76b | 19.08 ± 0.58 | 101.58 ± 0.80Bb | 12.75 ± 0.33 | 69.58 ± 0.97Bb | |||
| TT (57/81) | 74.99 ± 0.36 | 72.45 ± 0.36Bb | 84.11 ± 1.65 | 33.96 ± 0.80 | 28.02 ± 0.63a | 21.53 ± 0.34 | 106.68 ± 0.47Aa | 12.68 ± 0.11 | 68.66 ± 0.40Bb | |||
| Doe |
| C34197G | CC (693/1035) | 63.77 ± 0.14Ab | 65.77 ± 0.13Aa | 79.44 ± 0.23Aa | 32.15 ± 0.11b | 23.32 ± 0.15ab | 22.84 ± 0.17Aa | 99.54 ± 0.36Aa | 9.47 ± 0.04 | 63.68 ± 0.17Bb |
| CG (261/1035) | 64.66 ± 0.21Aa | 66.09 ± 0.18 Aa | 78.19 ± 0.36Aa | 32.86 ± 0.18a | 24.06 ± 0.28a | 22.99 ± 0.30Aa | 100.77 ± 0.67Aa | 9.52 ± 0.06 | 65.14 ± 0.28Aa | |||
| GG (81/1035) | 62.17 ± 0.35Bc | 63.39 ± 0.36 Bb | 75.33 ± 0.80Bb | 32.38 ± 0.20ab | 23.06 ± 0.45b | 21.09 ± 0.47Bb | 94.86 ± 0.70Bb | 9.49 ± 0.14 | 60.28 ± 0.35Cc | |||
| Buck |
| C34197G | CC (48/84) | 75.46 ± 0.43 | 73.25 ± 0.36 | 81.79 ± 2.28 | 32.33 ± 1.04b | 27.92 ± 0.86 | 20.48 ± 0.42 | 104.98 ± 0.72 | 12.71 ± 0.18 | 69.50 ± 0.56 |
| CG (36/84) | 74.94 ± 0.37 | 73.33 ± 0.69 | 87.39 ± 0.60 | 37.11 ± 0.24a | 25.81 ± 0.54 | 21.17 ± 0.42 | 105.10 ± 0.60 | 12.56 ± 0.11 | 69.50 ± 0.48 |
Different lowercase letters indicate significant differences (p < 0.05); different uppercase letters indicate highly significant differences (p < 0.01). Sample sizes were limited for some genotype groups (e.g.,
Different lowercase letters indicate significant differences (p < 0.05); different uppercase letters indicate highly significant differences (p < 0.01). Sample sizes were limited for some genotype groups (e.g.,
Effects of GMPS and SRC genotypes on slaughter performance in LCG
Genotype means ± standard error for slaughter performance traits are presented in Table 6. In bucks, the
Table 6. Liaoning cashmere goat slaughter performance: associations with
| Gender | Gene | Loci | Genotype (n/total) | Live weight before slaughter (kg) | Carcass weight (kg) | Net meat weight (kg) | Slaughter rate (%) | Net meat consumption rate (%) | Net carcass meat percentage (%) | Eye muscle area (cm²) | GR value (mm) | Back fat thickness (mm) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Buck |
| C31799T | CC (3/39) | 46.80 ± 0.50ab | 22.00 ± 0.61 | 17.40 ± 0.21 | 47.01 ± 0.30Bb | 37.18 ± 0.45 | 79.09 ± 0.31Aa | 19.65 ± 0.23 | 4.28 ± 0.15Bb | 1.06 ± 0.25b |
| CT (6/39) | 41.55 ± 0.25b | 21.75 ± 0.16 | 16.25 ± 0.11 | 52.34 ± 0.07Aa | 39.11 ± 0.04 | 74.71 ± 0.02Bb | 20.88 ± 1.11 | 7.94 ± 0.10Aa | 1.63 ± 0.21ab | |||
| TT (30/39) | 47.87 ± 0.94a | 24.81 ± 0.65 | 19.63 ± 0.60 | 51.72 ± 0.64ABa | 40.84 ± 0.67 | 78.86 ± 0.48Aa | 22.70 ± 0.88 | 6.21 ± 0.38Aab | 2.25 ± 0.18a | |||
| Doe |
| C31799T | CT (25/60) | 46.78 ± 1.20 | 22.78 ± 0.45 | 18.78 ± 0.37 | 48.91 ± 0.32b | 40.33 ± 0.33 | 82.44 ± 0.29 | 18.94 ± 0.46 | 6.98 ± 0.15b | 2.47 ± 0.09 |
| TT (35/60) | 45.67 ± 0.92 | 23.39 ± 0.56 | 19.17 ± 0.53 | 51.13 ± 0.38a | 41.84 ± 0.47 | 81.73 ± 0.36 | 19.88 ± 0.39 | 8.78 ± 0.30a | 3.11 ± 0.16 | |||
| Buck |
| C34197G | CC (30/60) | 49.35 ± 1.06Aa | 24.68 ± 0.60Aa | 20.33 ± 0.56Aa | 50.01 ± 0.47 | 41.14 ± 0.52 | 82.20 ± 0.27 | 19.59 ± 0.22Aa | 9.16 ± 0.25Aa | 3.04 ± 0.20 |
| CG (20/60) | 44.00 ± 0.87Bb | 22.20 ± 0.30ABb | 18.10 ± 0.34ABb | 50.63 ± 0.48 | 41.25 ± 0.55 | 81.43 ± 0.55 | 20.85 ± 0.61Aa | 7.13 ± 0.30Bb | 2.72 ± 0.07 | |||
| GG (10/60) | 40.75 ± 0.80Bb | 20.35 ± 0.35Bb | 16.85 ± 0.38Bb | 49.97 ± 0.13 | 41.32 ± 0.11 | 82.72 ± 0.44 | 16.43 ± 0.57Bb | 6.43 ± 0.16Bb | 2.50 ± 0.06 | |||
| Doe |
| C34197G | CC (24/32) | 47.22 ± 1.35 | 24.15 ± 0.92 | 18.97 ± 0.90 | 50.92 ± 0.76 | 39.78 ± 0.89 | 77.97 ± 0.80 | 20.02 ± 0.46b | 6.26 ± 0.48 | 2.31 ± 0.26 |
| CG (8/32) | 49.60 ± 1.28 | 26.05 ± 0.35 | 20.45 ± 0.36 | 52.64 ± 0.64 | 41.29 ± 0.35 | 78.47 ± 0.30 | 24.02 ± 1.66a | 7.16 ± 0.57 | 1.79 ± 0.04 |
Different lowercase letters indicate significant differences (p < 0.05); different uppercase letters indicate highly significant differences (p < 0.01). Sample sizes were limited for some genotype groups (e.g.,
Different lowercase letters indicate significant differences (p < 0.05); different uppercase letters indicate highly significant differences (p < 0.01). Sample sizes were limited for some genotype groups (e.g.,
Effects of GMPS and SRC gene variations on meat quality traits in LCG
Genotype means ± standard error for meat quality parameters are presented in Table 7. In bucks, the
Table 7. Analysis of the meat quality traits of the
| Gender | Gene | Loci | Genotype (n/total) | Meat color (L) | Meat color (a) | Meat color (b) | pH | Dry matter content (%) | Protein content (%) | Fat content (%) | Drip loss (%) | Cooked meat (%) | Shear force (N) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Buck |
| C31799T | CC (3/39) | 28.45 ± 0.12 | 18.54 ± 0.20Aa | 1.51 ± 0.04 | 6.02 ± 0.08 | 28.60 ± 0.25Aa | 21.00 ± 0.15 | 1.50 ± 0.07 | 1.12 ± 0.01Cc | 70.86 ± 0.09a | 70.85 ± 0.00Bb |
| CT (6/39) | 28.26 ± 0.21 | 13.06 ± 0.10Bc | 1.68 ± 0.02 | 6.04 ± 0.03 | 25.35 ± 0.11Bb | 21.81 ± 0.10 | 2.31 ± 0.08 | 1.52 ± 0.01Bb | 66.30 ± 0.04b | 82.34 ± 0.26Aa | |||
| TT (30/39) | 29.89 ± 0.18 | 14.72 ± 0.16Bb | 1.74 ± 0.04 | 6.10 ± 0.02 | 25.76 ± 0.15Bb | 21.38 ± 0.12 | 1.83 ± 0.09 | 1.73 ± 0.01Aa | 65.27 ± 0.43b | 74.66 ± 0.79ABab | |||
| Doe |
| C31799T | CT (25/60) | 30.33 ± 0.12 | 13.23 ± 0.03Bb | 1.87 ± 0.03 | 5.91 ± 0.01 | 29.48 ± 0.21 | 20.67 ± 0.07 | 2.29 ± 0.03 | 1.77 ± 0.01 | 69.65 ± 0.12 | 70.19 ± 0.59 |
| TT (35/60) | 29.56 ± 0.07 | 15.05 ± 0.05Aa | 1.81 ± 0.02 | 6.00 ± 0.01 | 26.73 ± 0.05 | 21.00 ± 0.04 | 2.34 ± 0.05 | 1.71 ± 0.01 | 67.65 ± 0.20 | 71.56 ± 0.24 | |||
| Buck |
| C34197G | CC (24/32) | 28.15 ± 0.17Bb | 14.66 ± 0.44 | 1.62 ± 0.02Bb | 6.11 ± 0.03b | 26.43 ± 0.35 | 20.86 ± 0.21Bb | 1.96 ± 0.08Aa | 1.58 ± 0.05 | 65.64 ± 1.03 | 76.26 ± 1.41a |
| CG (8/32) | 31.22 ± 0.02Aa | 14.02 ± 0.16 | 2.21 ± 0.22Aa | 6.25 ± 0.02a | 25.03 ± 0.00 | 22.50 ± 0.15Aa | 1.11 ± 0.08Bb | 1.79 ± 0.01 | 67.68 ± 0.99 | 69.40 ± 1.13b | |||
| Doe |
| C34197G | CC (30/60) | 28.69 ± 0.36Bc | 14.39 ± 0.23Aa | 1.80 ± 0.08 | 5.97 ± 0.03 | 26.97 ± 0.22Bb | 21.47 ± 0.20Aa | 2.66 ± 0.21Aa | 1.67 ± 0.06 | 69.65 ± 0.71 | 75.68 ± 1.30Aa |
| CG (20/60) | 31.62 ± 0.36Aa | 14.80 ± 0.37Aa | 1.77 ± 0.15 | 5.96 ± 0.02 | 30.15 ± 1.11Aa | 19.79 ± 0.20Bb | 1.61 ± 0.10Bb | 1.80 ± 0.05 | 67.37 ± 1.24 | 63.99 ± 2.13Bb | |||
| GG (10/60) | 29.98 ± 0.47ABb | 12.97 ± 0.12Bb | 2.07 ± 0.22 | 5.96 ± 0.01 | 26.04 ± 0.18Bb | 21.19 ± 0.06Aa | 2.70 ± 0.13Aa | 1.81 ± 0.01 | 67.23 ± 0.44 | 70.92 ± 1.90ABa |
Different lowercase letters indicate significant differences (p < 0.05); different uppercase letters indicate highly significant differences (p < 0.01). Sample sizes were limited for some genotype groups (e.g.,
Effects of GMPS and SRC gene variations on milk production traits in LCG
Milk composition parameters by genotype are shown in Table 8. For
Table 8. Association between
| Name | Gene | Loci | Genotype (n/total) | Fat | Crude protein | Lactose | Urea N | SnF | TS | Conductivity | Health index |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Doe |
| C31799T | CC (18/354) | 5.98 ± 0.30b | 4.43 ± 0.03b | 5.34 ± 0.02 | 38.63 ± 0.49b | 18.04 ± 0.23b | 10.23 ± 0.03 b | 16.48 ± 0.29Bb | 766.23 ± 6.45 |
| CT (108/354) | 7.11 ± 0.27a | 5.78 ± 0.36a | 5.14 ± 0.06 | 42.29 ± 1.09a | 19.74 ± 0.51a | 11.60 ± 0.35a | 19.10 ± 0.38Aa | 749.43 ± 6.36 | |||
| TT (228/354) | 7.14 ± 0.17a | 5.16 ± 0.12ab | 5.11 ± 0.05 | 39.92 ± 0.37ab | 18.62 ± 0.17ab | 10.85 ± 0.11ab | 18.38 ± 0.20Aa | 775.88 ± 7.52 | |||
| Doe |
| C34197G | CC (216/328) | 6.80 ± 0.16Aa | 5.14 ± 0.12ab | 5.14 ± 0.03Ab | 40.20 ± 0.40Aa | 18.76 ± 0.18Aa | 10.86 ± 0.11 | 18.03 ± 0.19Aa | 775.57 ± 4.21Bb |
| CG (88/328) | 7.60 ± 0.27Aa | 5.69 ± 0.28a | 4.74 ± 0.11Bc | 40.92 ± 0.39Aa | 19.10 ± 0.18Aa | 11.05 ± 0.23 | 19.09 ± 0.40Aa | 841.96 ± 18.89Aa | |||
| GG (24/328) | 5.05 ± 0.30Bb | 4.77 ± 0.05b | 5.46 ± 0.04Aa | 36.57 ± 0.44Bb | 17.07 ± 0.21Bb | 10.76 ± 0.05 | 16.05 ± 0.32Bb | 752.43 ± 4.61Bb |
Different lowercase letters indicate significant differences (p < 0.05); different uppercase letters indicate highly significant differences (p < 0.01). SnF = solids-not-fat; TS = total solids. Sample size was limited for
Effects of GMPS and SRC gene variants on lambing traits in LCG
Litter size means by genotype are presented in Table 9. No significant differences in the number of lambs per doe were detected across genotypes at either the
Table 9. Effects of
| Name | Gene | Loci | Genotype (n/total) | Number of lambs |
|---|---|---|---|---|
| Doe |
| C31799T | CT (145/325) | 1.38 ± 0.04 |
| TT (180/325) | 1.28 ± 0.03 | |||
| CC (215/325) | 1.35 ± 0.03 | |||
| Doe |
| C34197G | CG (85/325) | 1.29 ± 0.05 |
| GG (25/325) | 1.40 ± 0.10 | |||
| CT (145/325) | 1.38 ± 0.04 |
Relationships between fiber fineness and production traits in Liaoning cashmere goat
Pearson correlation coefficients between CF and other fiber and production traits are shown in Table 10. Fiber fineness exhibited highly significant positive correlations with shearing quantity (bucks r = 0.41, does r = 0.38; p < 0.01) and highly significant negative correlations with cashmere yield rate (bucks r = –0.90, does r = –0.93; p < 0.01) in both sexes.
Table 10. Relationships between fiber fineness and Liaoning cashmere goat production traits.
| Trait | Buck fineness (μm) | Doe fineness (μm) | Shearing quantity (g) Buck | Shearing quantity (g) Doe | Length (cm) Buck | Length (cm) Doe | Coefficient of variation in length Buck | Coefficient of variation in length Doe | Curl number Buck | Curl number Doe | Short fiber rate (%) Buck | Short fiber rate (%) Doe | Cashmere yield rate (%) Buck | Cashmere yield rate (%) Doe |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fineness (μm) | – | – | 0.41** | 0.38** | 0.09 | – | –0.06 | –0.32** | –0.09 | –0.14** | –0.26** | –0.15** | –0.90** | –0.93** |
| Shearing quantity (g) | 0.41** | 0.38** | – | – | –0.19 | 0.12** | 0.25* | –0.32** | 0.12 | –0.18** | 0.03 | –0.06* | –0.21* | –0.39** |
| Length (cm) | 0.09 | – | –0.19 | 0.12** | – | – | 0.07 | –0.47** | 0.38** | 0.04 | –0.26** | –0.37** | 0.00 | –0.35** |
| Coefficient of variation in length | –0.06 | –0.32** | 0.25* | –0.32** | 0.07 | –0.47** | – | – | 0.77** | 0.61** | 0.25* | 0.39** | 0.25* | 0.39** |
| Curl number | –0.09 | –0.14** | 0.12 | –0.18** | 0.38** | 0.04 | 0.77** | 0.61** | – | – | –0.25* | –0.15** | 0.21* | 0.12** |
| Short fiber rate (%) | –0.26** | –0.15** | 0.03 | –0.06* | –0.26** | –0.37** | 0.25* | 0.39** | –0.25* | –0.15** | – | – | 0.38** | 0.18** |
| Cashmere yield rate (%) | –0.90** | –0.93** | –0.21* | –0.39** | 0.00 | –0.35** | 0.25* | 0.39** | 0.21* | 0.12** | 0.38** | 0.18** | – | – |
Path analysis of CF on production traits
Direct and indirect path coefficients from production traits to CF are shown in Table 11. In does, the strongest direct negative effect was exerted by cashmere yield rate (–0.92), followed by coefficient of variation of length (0.10). In bucks, cashmere yield rate (–0.89) and shearing quantity (0.23) were the dominant direct contributors.
Table 11. Path analysis of cashmere fineness on Liaoning cashmere goat production traits.
| Gender | Independent variable | Correlation coefficient | Direct path coefficient | Indirect effect via Shearing quantity (g) | Indirect effect via Length (cm) | Indirect effect via Coefficient of variation in length | Indirect effect via Curl number | Indirect effect via Short fiber rate (%) | Indirect effect via Cashmere yield rate (%) |
|---|---|---|---|---|---|---|---|---|---|
| Doe | Shearing quantity (g) | 0.38 | 0.05 | – | 0.01 | –0.01 | –0.01 | 0.00 | –0.02 |
| Length (cm) | 0.34 | 0.06 | 0.01 | – | –0.03 | 0.00 | –0.02 | –0.02 | |
| Coefficient of variation in length | –0.32 | 0.10 | –0.03 | –0.05 | – | 0.01 | 0.06 | 0.04 | |
| Curl number | –0.14 | –0.03 | 0.01 | 0.00 | 0.00 | – | 0.01 | 0.00 | |
| Short fiber rate (%) | –0.15 | –0.02 | 0.00 | 0.01 | –0.01 | 0.00 | – | 0.00 | |
| Cashmere yield rate (%) | –0.93 | –0.92 | 0.36 | 0.32 | –0.36 | –0.11 | –0.17 | – | |
| Buck | Shearing quantity (g) | 0.41 | 0.23 | – | – | – | – | –0.04 | 0.06 |
| Short fiber rate (%) | –0.26 | 0.06 | –0.01 | – | – | – | – | 0.00 | |
| Cashmere yield rate (%) | –0.90 | –0.89 | –0.22 | – | – | – | –0.06 | – |
Stepwise regression analysis of cashmere traits and fineness
Multiple linear regression models with CF as the dependent variable are presented in Table 12. For bucks: CF = 0.001 × shearing quantity – 0.07 × cashmere yield rate + 20.553 (R² = 0.85). For does: CF = 0.0001 × shearing quantity – 0.079 × cashmere yield rate + 22.307 (R² = 0.86). Both models explained more than 85% of the variation in CF.
Table 12. Stepwise regression analysis of cashmere traits and fineness in Liaoning cashmere goat.
| Gender | Model | R² | Adjusted R² | Standard error of the estimate | F-value | p-value |
|---|---|---|---|---|---|---|
| Buck | CF = 22.360 – 0.07 CY | 0.80 | 0.80 | 0.51 | 306.39 | 0.00 |
| CF = 0.001 SQ – 0.07 CY + 20.553 | 0.85 | 0.85 | 0.44 | 218.42 | 0.00 | |
| Doe | CF = 22.450 – 0.078 CY | 0.86 | 0.86 | 0.47 | 6356.68 | 0.00 |
| CF = 0.0001 SQ – 0.079 CY + 22.307 | 0.86 | 0.86 | 0.47 | 3231.52 | 0.00 |
Association of CF with slaughter traits in LCG
Pearson correlation coefficients between CF and slaughter traits are shown in Table 13. Fineness exhibited highly significant negative correlations with several carcass merit indicators, including slaughter rate, net meat consumption rate, and net carcass meat percentage in both sexes (p < 0.01).
Table 13. Association between cashmere fineness and slaughter traits in Liaoning cashmere goat.
| Trait | Buck fineness (μm) | Doe fineness (μm) | Live weight before slaughter (kg) Buck | Live weight before slaughter (kg) Doe | Carcass weight (kg) Buck | Carcass weight (kg) Doe | Net meat weight (kg) Buck | Net meat weight (kg) Doe | Slaughter rate (%) Buck | Slaughter rate (%) Doe | Net meat consumption rate (%) Buck | Net meat consumption rate (%) Doe | Net carcass meat percentage (%) Buck | Net carcass meat percentage (%) Doe | Eye muscle area (cm²) Buck | Eye muscle area (cm²) Doe | GR value (mm) Buck | GR value (mm) Doe | Back fat thickness (mm) Buck | Back fat thickness (mm) Doe |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fineness (μm) | – | – | 0.11 | –0.20 | –0.08 | –0.33 | –0.13 | –0.39 | –0.477 | –0.48 | –0.48 | –0.54 | –0.32 | –0.57 | 0.24 | –0.48 | –0.18 | –0.09 | –0.31 | –0.28 |
| Live weight before slaughter (kg) | 0.11 | –0.20 | – | – | 0.92 | 0.90 | 0.88 | 0.90 | –0.197 | 0.21 | –0.02 | 0.45 | 0.31 | 0.70 | 0.31 | –0.19 | 0.32 | 0.27 | 0.37 | 0.61 |
| Carcass weight (kg) | –0.08 | –0.33 | 0.92 | 0.90 | – | – | 0.99 | 0.99 | 0.194 | 0.62 | 0.35 | 0.78 | 0.51 | 0.73 | 0.20 | 0.07 | 0.34 | 0.45 | 0.59 | 0.63 |
| Net meat weight (kg) | –0.13 | –0.39 | 0.88 | 0.90 | 0.99 | 0.99 | – | – | 0.269 | 0.59 | 0.45 | 0.80 | 0.62 | 0.82 | 0.13 | 0.05 | 0.30 | 0.46 | 0.61 | 0.68 |
| Slaughter rate (%) | –0.477 | –0.48 | –0.197 | 0.21 | 0.194 | 0.62 | 0.269 | 0.59 | – | – | 0.94 | 0.919 | 0.51 | 0.381 | –0.27 | 0.504 | –0.03 | 0.544 | 0.52 | 0.28 |
| Net meat consumption rate (%) | –0.48 | –0.54 | –0.02 | 0.45 | 0.35 | 0.78 | 0.45 | 0.80 | 0.94 | 0.919 | – | – | 0.76 | 0.71 | –0.33 | 0.38 | –0.05 | 0.59 | 0.53 | 0.51 |
| Net carcass meat percentage (%) | –0.32 | –0.57 | 0.31 | 0.70 | 0.51 | 0.73 | 0.62 | 0.82 | 0.51 | 0.381 | 0.76 | 0.71 | – | – | –0.33 | 0.00 | –0.07 | 0.36 | 0.36 | 0.67 |
| Eye muscle area (cm²) | 0.24 | –0.48 | 0.31 | –0.19 | 0.20 | 0.07 | 0.13 | 0.05 | –0.27 | 0.504 | –0.33 | 0.38 | –0.33 | 0.00 | – | – | 0.15 | –0.28 | –0.01 | –0.24 |
| GR value (mm) | –0.18 | –0.09 | 0.32 | 0.27 | 0.34 | 0.45 | 0.30 | 0.46 | –0.03 | 0.544 | –0.05 | 0.59 | –0.07 | 0.38 | 0.15 | –0.28 | – | – | 0.60 | 0.32 |
| Back fat thickness (mm) | –0.31 | –0.28 | 0.37 | 0.61 | 0.59 | 0.63 | 0.61 | 0.68 | 0.52 | 0.28 | 0.53 | 0.51 | 0.36 | 0.67 | –0.01 | –0.24 | 0.60 | 0.32 | – | – |
** p < 0.01;
* p < 0.05; no symbol = p > 0.05.
Path analysis of slaughter traits on CF
Direct and indirect path coefficients from slaughter traits to CF are presented in Table 14. In bucks, net meat weight exerted the strongest direct positive effect on fineness (31.82). In does, the slaughter rate was the primary direct negative contributor (–1.05).
Table 14. Path analysis of Liaoning cashmere goat slaughter traits on cashmere fineness.
| Gender | Independent variable | Correlation coefficient | Direct path coefficient | Indirect action (path coefficient) Net meat weight (kg) | Indirect action (path coefficient) Slaughter rate (%) | Indirect action (path coefficient) Net meat consumption rate (%) | Indirect action (path coefficient) Net carcass meat percentage (%) | Indirect action (path coefficient) Eye muscle area (cm²) | Indirect action (path coefficient) Back fat thickness (mm) |
|---|---|---|---|---|---|---|---|---|---|
| Buck | Net meat weight (kg) | –0.39 | 31.82 | – | 18.84 | 25.36 | 26.00 | 1.50 | – |
| Slaughter rate (%) | –0.40 | 10.52 | 6.23 | – | 9.67 | 4.01 | 5.30 | – | |
| Net meat consumption rate (%) | –0.54 | –29.92 | –23.84 | –27.49 | – | –21.36 | –11.28 | – | |
| Net carcass meat percentage (%) | –0.57 | 5.89 | 4.81 | 2.24 | 4.21 | – | 0.02 | – | |
| Eye muscle area (cm²) | –0.48 | –0.10 | 0.00 | –0.05 | –0.04 | 0.00 | – | – | |
| Doe | Slaughter rate (%) | –0.48 | –1.05 | – | – | – | –0.53 | – | –0.54 |
| Net carcass meat percentage (%) | –0.32 | –0.52 | – | –0.26 | – | – | – | –0.19 | |
| Back fat thickness (mm) | –0.31 | 0.04 | – | 0.02 | – | 0.01 | – | – |
Stepwise regression model of slaughter traits for CF
Optimal multiple regression equations for CF based on slaughter traits are shown in Table 15. For does: CF = 27.558 – 0.217 × slaughter rate (R² = 0.23). For bucks: CF = 36.723 – 0.230 × net carcass meat percentage – 0.118 × eye muscle area (R² = 0.55).
Table 15. Stepwise regression model of slaughter traits for fineness of cashmere in Liaoning cashmere goat.
| Gender | Model | R² | Adjusted R² | Standard error of the estimate | F-value | p-value |
|---|---|---|---|---|---|---|
| Buck | CF = 34.164 – 0.230 CP | 0.33 | 0.31 | 0.92 | 18.02 | 0.00 |
| CF = 36.723 – 0.230 CP – 0.118 EM | 0.55 | 0.53 | 0.77 | 22.08 | 0.00 | |
| Doe | CF = 27.558 – 0.217 DP | 0.23 | 0.21 | 0.98 | 17.08 | 0.00 |
CF = Cashmere fineness; CP = Net carcass meat percentage; EM = Eye muscle area; DP = Slaughter rate.
Association of GMPS and SRC haplotypes with production traits
Nine haplotype combinations were inferred from the two SNP loci using SHEsis software (Table 16). Bucks displayed six combinations: CCCC, CCCG, CTCC, CTCG, TTCC, TTCG. Does displayed seven combinations: CCGG, CTCC, CTCG, CTGG, TTCC, TTCG, TTGG.
Table 16. Haplotype combination analysis of the
| Haplotype combination | CC | CG | GG |
|---|---|---|---|
| CC | CCCC | CCCG | CCGG |
| CT | CTCC | CTCG | CTGG |
| TT | TTCC | TTCG | TTGG |
Haplotype effects on cashmere production performance
Haplotype means ± standard error for cashmere traits are presented in Table 17. In bucks, the CCCC combination was associated with the finest cashmere fibers. In does, the TTGG combination outperformed the other haplotypes in fineness, coefficient of variation of length, and curl number.
Table 17. Haplotype effects of
| Name | Haplotype | Shearing quantity (g) | Fineness (μm) | Length (cm) | Coefficient of variation in length (%) | Curl number | Short fiber rate (%) | Cashmere yield rate (%) |
|---|---|---|---|---|---|---|---|---|
| Buck | CCCC (4/80) | 1700.00 ± 30.15Cc | 15.92 ± 21.31Bc | 82.80 ± 5.30Cc | 50.98 ± 36.70ab | 9.40 ± 29.40 | 0.09 ± 32.00Bb | 0.75 ± 28.60ab |
| CCCG (4/80) | 2000.00 ± 15.05BCb | 17.57 ± 50.10Aa | 154.80 ± 3.06Aa | 42.22 ± 46.80b | 8.70 ± 52.60 | 0.09 ± 49.03Bb | 0.71 ± 47.62 ab | |
| CTCC (16/80) | 1925.00 ± 62.04BCbc | 17.13 ± 62.04ABab | 100.05 ± 3.03BCbc | 52.57 ± 62.04ab | 8.43 ± 62.04 | 0.08 ± 62.04Bb | 0.68 ± 62.04b | |
| CTCG (4/80) | 2500.00 ± 20.50Aa | 17.25 ± 35.20ABab | 113.80 ± 2.45Bb | 55.99 ± 42.60ab | 7.60 ± 50.70 | 0.17 ± 36.50ABab | 0.67 ± 41.20b | |
| TTCC (24/80) | 2100.00 ± 41.87Bb | 16.80 ± 41.87ABabc | 86.97 ± 3.42Cc | 57.22 ± 41.87a | 7.52 ± 41.87 | 0.17 ± 41.87ABab | 0.71 ± 41.87ab | |
| TTCG (28/80) | 2092.86 ± 62.08Bb | 16.34 ± 62.08ABbc | 88.09 ± 3.43Cc | 58.96 ± 62.08a | 6.64 ± 62.08 | 0.31 ± 62.08Aa | 0.82 ± 62.08a | |
| Doe | CCGG (15/1035) | 2350.00 ± 15.20Aa | 18.09 ± 0.20Aa | 61.60 ± 2.80Cc | 25.39 ± 0.90Cd | 6.78 ± 0.30 | 0.21 ± 0.02Aa | 0.54 ± 0.02Cd |
| CTCC (270/1035) | 1783.33 ± 17.34Ccd | 16.87 ± 0.08BCb | 100. ± 2.02Bb | 52.80 ± 1.02Bbc | 6.73 ± 0.23ABb | 0.15 ± 0.01Ab | 0.72 ± 0.01Bc | |
| CTCG (135/1035) | 1700.00 ± 20.86Cd | 16.14 ± 0.11DEc | 97.9 ± 2.95Bb | 53.66 ± 1.10Bbc | 6.57 ± 0.42ABb | 0.20 ± 0.01Aab | 0.79 ± 0.01Bb | |
| CTGG (30/1035) | 1950.00 ± 37.14Bb | 16.92 ± 0.07Bb | 135. ± 0.84Aa | 56.82 ± 0.76ABab | 4.65 ± 0.86Bc | 0.21 ± 0.02Aa | 0.75 ± 0.01Bbc | |
| TTCC (405/1035) | 1838.89 ± 15.78BCbc | 16.71 ± 0.06BCDb | 96.89 ± 1.60Bb | 55.30 ± 0.57ABbc | 6.55 ± 0.20ABb | 0.20 ± 0.01Aab | 0.74 ± 0.01Bbc | |
| TTCG (135/1035) | 1766.67 ± 19.95Ccd | 16.22 ± 0.09CDc | 97.64 ± 1.63Bb | 51.02 ± 0.67Bc | 8.58 ± 0.16Aa | 0.04 ± 0.01Bc | 0.78 ± 0.01Bbc | |
| TTGG (45/1035) | 1550.00 ± 30.77De | 15.53 ± 0.16Ed | 87.00 ± 3.17Bb | 61.24 ± 1.27Aa | 8.30 ± 0.16Aa | 0.20 ± 0.01Aab | 0.88 ± 0.01Ca |
Different lowercase letters indicate significant differences (p < 0.05); different uppercase letters indicate highly significant differences (p < 0.01). Sample sizes were limited for several haplotypes (n < 20).
Haplotype effects on slaughter traits
Haplotype means ± standard error for slaughter performance traits are presented in Table 18. In bucks, TTCC was the superior haplotype for live weight before slaughter, carcass weight, and back fat thickness. In does, CTCC demonstrated consistent superiority across multiple slaughter traits.
Table 18. Effects of haplotype on slaughter traits in Liaoning cashmere goat (mean ± standard error).
| Name | Haplotype | Live weight before slaughter (kg) | Carcass weight (kg) | Net meat weight (kg) | Slaughter rate (%) | Net meat consumption rate (%) | Net carcass meat percentage (%) | Eye muscle area (cm²) | GR value (mm) | Back fat thickness (mm) |
|---|---|---|---|---|---|---|---|---|---|---|
| Buck | CCCC (4/32) | 46.80 ± 0.30ABab | 22.00 ± 0.25b | 17.40 ± 0.06ab | 47.01 ± 0.11Bb | 37.18 ± 0.20 | 79.09 ± 0.25ABa | 19.65 ± 0.70b | 4.28 ± 0.62Bb | 1.06 ± 0.12Bb |
| CTCC (8/32) | 41.55 ± 0.21Bb | 21.75 ± 0.13b | 16.25 ± 0.09b | 52.34 ± 0.06Aa | 39.11 ± 0.03 | 74.71 ± 0.02Bb | 20.88 ± 0.94ab | 7.94 ± 0.09Aa | 1.63 ± 0.18Bb | |
| TTCC (12/32) | 51.13 ± 2.07Aa | 26.47 ± 1.61a | 21.30 ± 1.53a | 51.28 ± 1.36ABa | 41.09 ± 1.72 | 79.76 ± 1.31Aa | 19.57 ± 0.66b | 5.80 ± 0.82ABab | 3.18 ± 0.36Aa | |
| TTCG (8/32) | 49.60 ± 1.29Aa | 26.05 ± 0.36a | 20.45 ± 0.36a | 52.64 ± 0.64Aa | 41.29 ± 0.35 | 78.47 ± 0.30ABa | 24.02 ± 1.66a | 7.16 ± 0.57ABa | 1.79 ± 0.04Bb | |
| Doe | CTCC (6/72) | 57.50 ± 0.53Aa | 26.20 ± 0.45Aa | 21.20 ± 0.26Aa | 45.57 ± 0.05Bc | 36.87 ± 0.15Bb | 80.92 ± 0.35ABbc | 20.93 ± 0.46Aab | 6.39 ± 0.09Cc | 1.65 ± 0.18Bc |
| CTCG (12/72) | 47.45 ± 0.59Bb | 23.50 ± 0.27ABb | 19.50 ± 0.33ABa | 49.53 ± 0.04Ab | 41.07 ± 0.19Aa | 82.92 ± 0.45Aa | 20.45 ± 0.35Aab | 7.84 ± 0.07Bb | 2.85 ± 0.12Aab | |
| CTGG (12/72) | 40.75 ± 0.80Cc | 20.35 ± 0.35Cc | 16.85 ± 0.38Bb | 49.97 ± 0.13Ab | 41.32 ± 0.11Aa | 82.71 ± 0.44Aa | 16.43 ± 0.56Bc | 6.43 ± 0.16Cc | 2.50 ± 0.06ABb | |
| TTCC (30/72) | 47.72 ± 1.03Bb | 24.38 ± 0.71Aba | 20.16 ± 0.67Aa | 50.89 ± 0.40Aab | 41.98 ± 0.49Aa | 82.45 ± 0.31Aab | 19.33 ± 0.24Ab | 9.72 ± 0.17Aa | 3.32 ± 0.21Aa | |
| TTCG (12/72) | 40.55 ± 0.80Cc | 20.90 ± 0.60BCc | 16.70 ± 0.12Bb | 51.73 ± 0.87Aa | 41.43 ± 1.11Aa | 79.93 ± 0.81Bc | 21.25 ± 1.18Aa | 6.43 ± 0.53Cc | 2.58 ± 0.62Ab |
Different lowercase letters indicate significant differences (p < 0.05); different uppercase letters indicate highly significant differences (p < 0.01). Sample sizes were limited for several haplotypes (n < 20).
DISCUSSION
Population genetic characteristics of the C31799T and C34197G loci
This deviation is likely attributable to multiple factors. Primarily, the long-term, intensive artificial selection practiced to improve CF and growth performance has likely led to directed changes in allele frequencies and the evolution of the population’s genetic structure [17]. Additionally, factors such as the limited size of the breeding population, skewed sex ratios, or a certain degree of inbreeding may have exacerbated this deviation [18]. In our statistical analyses, we accounted for these potential confounding factors by incorporating pedigree information (to control for kinship), and by including sex and age as fixed effects in a general linear model. This approach was implemented to mitigate the interference of these factors on the detected association signals.
Potential biological roles of GMPS in CF
Potential biological roles of SRC in CF and slaughter performance
On the other hand, the
Pleiotropic effects and sex-specific genetic regulation of production traits
Comparison with genetic studies in other cashmere goat breeds
The effects of
Correlations among cashmere production and slaughter traits
The present study demonstrates a significant correlation between cashmere yield and cashmere length, with the greatest indirect effect coefficient being 0.318. Through research conducted by Feng
Implications for molecular breeding and future research directions
This study reveals statistically significant associations rather than direct causal relationships. Nonetheless, the identified superior genotypes and haplotype combinations provide valuable pre-selection markers for molecular breeding in LCGs. The observed negative correlation between CF and slaughter traits may stem from pleiotropy, tight genetic linkage, or indirect effects of physiological resource allocation, constituting a key focus for subsequent functional studies to elucidate the specific molecular interactions. In practical breeding, where the core objectives are to consistently reduce fiber diameter, each micrometer reduction can significantly enhance market value, while maintaining high yield, we propose a MAS strategy involving: 1) the establishment of sex-specific selection protocols, and 2) the development of criteria for early genetic evaluation and retention of replacement breeding stock based on the significant effects of
CONCLUSION
This study is the first to establish significant associations between polymorphisms in
The superior genotypes (
The investigation was performed on a large, well-managed nucleus herd (n = 1 160) with comprehensive phenotyping of cashmere production traits, body size measurements, slaughter performance, meat quality attributes, milk composition, and lambing rate. Rigorous statistical approaches, including general linear mixed models with pedigree-based relatedness control, sex-stratified haplotype inference, path analysis, and stepwise regression, provided robust adjustment for kinship, age, and sex effects. The combination of functional biological knowledge of
The associations were detected within a single nucleus population and therefore require independent validation in additional LCG herds and, ideally, in other cashmere goat breeds to confirm wider applicability. The identified SNPs are intronic; their exact functional consequences (e.g., on splicing efficiency, regulatory element activity, or gene expression) remain to be elucidated. Although major environmental and management factors were standardized and controlled, residual population stratification or unmeasured gene–environment interactions cannot be completely ruled out.
Priority should be assigned to (1) functional validation of the candidate SNPs through
By identifying
DATA AVAILABILITY
All the generated data are included in the manuscript.
AUTHORS’ CONTRIBUTIONS
QY: Formal data analysis, interpretation and drafted and revised the manuscript. RD and QZ: Study design and experimental implementation, data collection, and final manuscript revision. SL, LK, YZ, HL, HM, WH, WL, HW, XK, DL, YG, and JL: Collected and pretreated experimental samples (core step of research data collection), verified experimental data, and final manuscript review. ZW: Conceived and designed the study, developed core research methodologies, supervised the entire process of experiments and data analysis, and revised and finalized the manuscript. All authors have read and approved the final version of the manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
We thank LCG Breeding Center for providing experimental animals and related equipment to help this study. This research was funded by National Natural Science Foundation of China (NO. 32272836).
REFERENCES
- Wang Z, Liu R. Growth regularity of cashmere goats in Liaoning province. Today Anim Husb Vet 2020;36(06):53+61. [Google Scholar] | [Crossref]
- Fan Z. The Research of Conservation and Utility of Liaoning Cashmere Goats Resource [D] 2013. [Google Scholar] | [Crossref]
- Zhang X, Zhang Q. Analysis of Factors Affecting the Down Production Performance of LCGs. Contemp Anim Husb 2024;(07):39-41. [Google Scholar] | [Crossref]
- Khunsriraksakul C, McGuire D, Sauteraud R, Chen F, Yang L, Wang L. Integrating 3D genomic and epigenomic data to enhance target gene discovery and drug repurposing in transcriptome-wide association studies. Nat Commun 2022;13(1):3258. [Google Scholar] | [Crossref]
- Guldbrandsen HO, Staehr C, Iversen NK, Postnov DD, Matchkov VV. Does SRC kinase-mediated vasoconstriction impair penumbral reperfusion?. Stroke 2021;52(6):e250-e258. [Google Scholar] | [Crossref]
- Byeon SE, Yi YS, Oh J, Yoo BC, Hong S, Cho JY. The role of SRC kinase in macrophage-mediated inflammatory responses. Mediators Inflamm 2012;2012:512926. [Google Scholar] | [Crossref]
- Hirst M, Haliday E, Nakamura J, Lou L. Human GMP synthetase. Protein purification, cloning, and functional expression of cDNA. J Biol Chem 1994;269(38):23830-23837. [Google Scholar] | [Crossref]
- Zalkin H. GMP synthetase. Methods Enzymol 1985;113:273-278. [Google Scholar] | [Crossref]
- Faesen AC, Dirac AM, Shanmugham A, Ovaa H, Perrakis A, Sixma TK. Mechanism of USP7/HAUSP activation by its C-terminal ubiquitin-like domain and allosteric regulation by GMP-synthetase. Mol Cell 2011;44(1):147-159. [Google Scholar] | [Crossref]
- Reddy BA, van der Knaap JA, Bot AG, Mohd-Sarip A, Dekkers DH, Timmermans MA. Nucleotide biosynthetic enzyme GMP synthase is a TRIM21-controlled relay of p53 stabilization. Mol Cell 2014;53(3):458-470. [Google Scholar] | [Crossref]
- van der Knaap JA, Kozhevnikova E, Langenberg K, Moshkin YM, Verrijzer CP. Biosynthetic enzyme GMP synthetase cooperates with ubiquitin-specific protease 7 in transcriptional regulation of ecdysteroid target genes. Mol Cell Biol 2010;30(3):736-744. [Google Scholar] | [Crossref]
- van der Knaap JA, Kumar BR, Moshkin YM, Langenberg K, Krijgsveld J, Heck AJ. GMP synthetase stimulates histone H2B deubiquitylation by the epigenetic silencer USP7. Mol Cell 2005;17(5):695-707. [Google Scholar] | [Crossref]
- Bianchi-Smiraglia A, Wawrzyniak JA, Bagati A, Marvin EK, Ackroyd J, Moparthy S. Pharmacological targeting of guanosine monophosphate synthase suppresses melanoma cell invasion and tumorigenicity. Cell Death Differ 2015;22(11):1858-1864. [Google Scholar] | [Crossref]
- Holzer K, Drucker E, Roessler S, Dauch D, Heinzmann F, Waldburger N. Proteomic Analysis Reveals GMP Synthetase as p53 Repression Target in Liver Cancer. Am J Pathol 2017;187(2):228-235. [Google Scholar] | [Crossref]
- Descotes F, Jézéquel P, Spyratos F, Campion L, Grenot C, Lerebours F. Identification of potential prognostic biomarkers for node-negative breast tumours by proteomic analysis:a multicentric 2004 national PHRC study. Int J Oncol 2012;41(1):92-104. [Google Scholar] | [Crossref]
- Massière F, Badet-Denisot MA. The mechanism of glutamine-dependent amidotransferases. Cell Mol Life Sci 1998;54(3):205-222. [Google Scholar] | [Crossref]
- Lachance J. Detecting selection-induced departures from Hardy-Weinberg proportions. Genet Sel Evol 2009;41(1):15. [Google Scholar] | [Crossref]
- Hattemer HH. Genetic distance between populations :Part 3:Wahlund's principle as related to genetic distance and an application. Theor Appl Genet 1982;62(3):219-223. [Google Scholar] | [Crossref]
- Long H, Cameron S, Yu L, Rao Y. De novo GMP synthesis is required for axon guidance in
Drosophila . Genetics 2006;172(3):1633-1642. [Google Scholar] | [Crossref] - Rodriguez-Suarez R, Xu D, Veillette K, Davison J, Sillaots S, Kauffman S. Mechanism-of-action determination of GMP synthase inhibitors and target validation in
Candida albicans andAspergillus fumigatus . Chem Biol 2007;14(10):1163-1175. [Google Scholar] | [Crossref] - Tojo S, Hirooka K, Fujita Y. Expression of kinA and kinB of
Bacillus subtilis , necessary for sporulation initiation, is under positive stringent transcription control. J Bacteriol 2013;195(8):1656-1665. [Google Scholar] | [Crossref] - Ya-Qin Y, Huai-Yan C, Yuan-Yuan L, Lu G. Transcriptome analysis in fetal lungs of SRC1/SRC2 double-knockout mice. Acta Physiologica Sinica 2022;74(02):246-254. [Google Scholar] | [Crossref]
- Yoo JY, Shin H, Kim TH, Choi WS, Ferguson SD, Fazleabas AT. CRISPLD2 is a target of progesterone receptor and its expression is decreased in women with endometriosis. PLoS One 2014;9(6):e100481. [Google Scholar] | [Crossref]
- Collins JJ, Kuypers E, Nitsos I, Jane Pillow J, Polglase GR, Kemp MW. LPS-induced chorioamnionitis and antenatal corticosteroids modulate Shh signaling in the ovine fetal lung. Am J Physiol Lung Cell Mol Physiol 2012;303(9):L778-787. [Google Scholar] | [Crossref]
- Chinenov Y, Gupte R, Dobrovolna J, Flammer JR, Liu B, Michelassi FE. Role of transcriptional coregulator GRIP1 in the anti-inflammatory actions of glucocorticoids. Proc Natl Acad Sci U S A 2012;109(29):11776-11781. [Google Scholar] | [Crossref]
- DI R, He X, Guan W, Pu Y, Zhao Q, Ma Y. Initial analysis of genetic diversity status and phylogenetic relationship of Chinese cashmere goats breeds using microsatellite makers. J Northwest A F Univ (Nat Sci Ed) 2008;(04):15-21. [Google Scholar] | [Crossref]
- Bai J, Liu X, Liu L, Song Y, Zhang X, Song X. Correlation Analysis of Notch2 Gene Polymorphism with Cashmere and Growth Traits in Shanbei White Cashmere Goats. Chin J Anim Sci 2023;50(09):3630-3640. [Google Scholar] | [Crossref]
- Bin L. Selection, Identification and SNP Analysis of Correlation Genes with Cashmere Growth on Cashmere Goats [D]. Inner Mongolia Agricultural University 2012. [Google Scholar] | [Crossref]
- Liu H, Wang C, Zhu X, Jia Z. Effect of KAP8.1 Gene Polymorphism on Cashmere Traits in Inner Mongolia Cashmere Goats;proceedings of the National Sheep Production and Academic Symposium of the Sheep Branch, Chinese Association of Animal Science and Veterinary Medicine, Yinchuan, Ningxia, China, F, 2010 [C]. [Google Scholar] | [Crossref]
- Wu C, Ma S, Zhao B, Qin C, Wu Y, Di J. Drivers of plateau adaptability in cashmere goats revealed by genomic and transcriptomic analyses. BMC Genomics 2023;24(1):428. [Google Scholar] | [Crossref]
- Feng P, Ji Z, Gao Y, Yan H, Qu L. Correlation and Regression Analysis of Body Measurements, Body Weight and Cashmere Yield in Shanbei White Cashmere Goats. Heilongjiang Anim Sci Vet Med 2011;(01):42-44. [Google Scholar] | [Crossref]
- He X, Zhang T, Zhang Y, Geng S. Multiple statistical analysis of the effect of body measurement, wool trait and economic trait on cashmere goat. J Northwest A F Univ (Nat Sci Ed) 2004;(01):85-88+92. [Google Scholar] | [Crossref]
- Li L, You W, Wang X, Liu W, Li R, Li T. Correlation Analysis of Cashmere Yield with Body Size Traits, Body Weight, Cashmere Length, and Cashmere Fineness in Shanbei White Cashmere Goats. Chin J Anim Sci 2025;61(05):138-144. [Google Scholar] | [Crossref]
- Shi L, Cao D, Zhang Q, Zhu H, Jing D, Qu L. Analysis of variation coefficient of fiber fineness and length of Shanbei white cashmere goat. Heilongjiang Anim Sci Vet Med 2022;(12):43-45+49. [Google Scholar] | [Crossref]
- Zhang F, Liu W, Hu D, Zhang X. 2009. Analysis on Correlation of Phenotypic Economic Traits of Nanjiang Cashmere Goat. J. Domestic Anim Ecol 30;(01):26-28+33. [Google Scholar] | [Crossref]